






Авторы: Резникова Ж.И., Рябко Б.Я.
Показано, что время передачи информации у муравьев пропорционально количеству информации в сообщении. Для выяснения этого разработан эксперимент, в ходе которого муравьи для получения пищи должны были передавать друг другу известное экспериментатору количество информации. Установлено также, что муравьи, передавая «текст», способны использовать простейшие закономерности для его сжатия.
§1. Введение
Первоначально теория информации развивалась как математическая теория связи [ 1 ]. Но сразу же было ясно, что понятия энтропии и информации, сформулированные в этой теории, должны играть важную роль в изучении не только технических, но и других систем коммуникации. В частности, была оценена энтропия текстов на различных естественных языках человека (см. обзор в [ 2 ]). В данной работе показано, что понятия теории информации могут применяться и при анализе систем коммуникации животных, причем позволяют не только оценить те или иные количественные характеристики их сигнальной деятельности, но и могут служить основой опытов, позволяющих ответить на ключевой вопрос о наличии или отсутствии у них «языка».
Здесь необходимо сказать несколько слов о самом понятии «язык животных». В настоящее время достаточно ясного и общепринятого определения языка не существует, более того, по-видимому, не существует даже языка (парадигмы), на котором можно было бы определить понятие «язык животных». Обсуждение этих вопросов увело бы нас слишком далеко, отметим только, что большинство исследователей считают основным свойством и критерием языка возможность составления большого (потенциально бесконечного) числа осмысленных текстов (фраз) [ 3 ]. Считаем, что язык должен обладать еще одним свойством — длина сообщения должна быть приблизительно пропорциональна количеству информации в этом сообщении (этим свойством обладают языки человека [ 2 ]). В дальнейшем под языком будем понимать систему коммуникаций, обладающую этими двумя свойствами.
Отсутствие общепринятого определения языка животных свидетельствует не о слабом внимании этологов к, этим проблемам — наоборот, в настоящее время в этологии активно изучаются системы коммуникации животных и связанные с ними интеллектуальные способности [3, 4 ] . Естественно, что наиболее интесивно исследуются высоко социальные виды: обезьяны, дельфины, пингвины, пчелы, муравьи и т. д. Хотя многие этологи и зоологи чувствуют, что для сложной совместной деятельности социальных животных (и людей) им необходим язык (это, по-видимому, впервые сформулировано в легенде о Вавилонской башне), до сих пор, несмотря на значительное число публикаций (см. обзоры [ 3, 5 ]), ни для одного вида животных не доказано наличие развитого «языка», возможно,, за исключением дельфинов [ 6 ]. В частности, классический результат К. Фриша, доказавшего существование языка пчел [ 7 ], в последние годы оспаривается многими исследователями [ 5, 8 ]. Обезьян в неволе удалось научить началам жестового языка глухонемых и тем самым показать, чта им доступно использование знаково-понятийного аппарата [ 9 ]. Однако вопрос о существовании у них естественного языка остается открытым.
Сигнальная деятельность муравьев также привлекала внимание многих исследователей. Еще в прошлом веке [ 10 ] было высказано предположение о наличии у муравьев языка жестов, связанного с тонкими движениями антенн (тактильный или антеннальный код). К настоящему времени можно считать доказанным сам факт передачи информации с помощью тактильного кода [ 11 ]. Однако попытки непосредственной расшифровки сигналов муравьев и других животных до сих пор не увенчались успехом. Это объясняется значительными трудностями — техническими, связанными с большой подвижностью, сложностью наблюдения в естественной обстановке, и логическими, связанными с объективным выделением единиц коммуникации, с необходимостью выяснения побудительных причин сигнализаций и т. д.
Предлагаем принципиально иной подход к изучению языка муравьев, основанный на идеях теории информации. Суть подхода в следующем: в эксперименте создавалась ситуация, в которой муравьи для получения пищи должны были передавать друг другу известное экспериментатору количество информации. Эта информация — сведения о последовательности поворотов на пути к кормушке. Количество поворотов (развилок) менялось от 1 до 6 в разных опытах. При этом фиксировалось время антеннального контакта муравьев друг с другом, т. е. время, затрачиваемое ими на передачу сведений о последовательности поворотов.
Наши опыты показали, что время передачи сообщения муравьями пропорционально числу поворотов, т. е. количеству информации (по Шеннону) в этом сообщении. Это позволило оценить скорость передачи информации. Оказалось, что у разных видов муравьев скорость передачи информации различна; некоторые виды вообще неспособны к языковому общению. Эти результаты излагаются в §3.
Разработана также серия тестов, позволяющих оценить некоторые интеллектуальные способности муравьев, связанные с возможностью использования закономерностей «текста» для его «сжатия». Точнее, муравьям предлагалась «закономерная» последовательность поворотов на пути к кормушке, например такая, в которой повороты налево и направо чередуются. Оказалось, что муравьи затрачивают значительно меньше времени на передачу сведений о такой последовательности поворотов, чем о случайной последовательности той же длины, т. е. муравьи способны использовать простейшие закономерности текста для его «сжатия».
Выражаясь несколько неточно, можно сказать, что опыты из §3 основаны на вероятностном подходе к понятию «количество информации», восходящем к К. Шеннону [ 1 ], а опыты §4 —на алгоритмическом подходе, восходящем к А. Н. Колмогорову [ 12 ]. Из опытов, описанных в §3, следует, что число реальных сообщений в «языке» муравьев равно 27-1=123 (максимальное число поворотов на пути к кормушке равно 6, каждый поворот ведет налево или направо).
Из опытов §4 следует, что, по-видимому, число потенциально возможных сообщений (или фраз, текстов) в «языке» муравьев значительно больше, так как они легко находят специальные короткие фразы, кодирующие «закономерные» последовательности поворотов.
Таким образом, показано, что в системе коммуникации муравьев содержится довольно много потенциально возможных сообщений, а время передачи сообщений пропорционально количеству информации в них. Нам кажется, что предложенный здесь подход может быть применен для изучения языка и интеллектуальных способностей и других социальных животных. Кроме того, он дает возможность количественного сравнения «языков » различных видов животных.
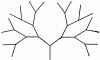
Схема лабиринта — «бинарного дерева» с четырьмя развилками
§2. Методика эксперимента
Исследовались 3 вида муравьев (Formica polyctena, ![]() F. sanguinea, Camponotus saxatilis), содержащиеся в искусственных гнездах на лабораторных аренах. Муравьи получали пищу развЗ дня и только в лабиринте. Лабиринт был смонтирован в виде «бинарного дерева». Каждый лист заканчивался кормушкой, но только одна из них содержала сахарный сироп, остальные были пусты. В простейшем случае установка состояла из дерева с двумя листьями и соответственно с двумя кормушками, в одной из которых был сироп. В такой ситуации муравей-разведчик должен был передать муравьям-фуражирам 1 бит информации — идти надо либо направо, либо налево. В ходе опыта число узлов (развилок) на одной ветви возрастало до 6 (рисунок). При этом количество информации (в битах), предположительно передаваемой разведчиками, было равно числу узлов дерева (поворотов) на пути к кормушке.
F. sanguinea, Camponotus saxatilis), содержащиеся в искусственных гнездах на лабораторных аренах. Муравьи получали пищу развЗ дня и только в лабиринте. Лабиринт был смонтирован в виде «бинарного дерева». Каждый лист заканчивался кормушкой, но только одна из них содержала сахарный сироп, остальные были пусты. В простейшем случае установка состояла из дерева с двумя листьями и соответственно с двумя кормушками, в одной из которых был сироп. В такой ситуации муравей-разведчик должен был передать муравьям-фуражирам 1 бит информации — идти надо либо направо, либо налево. В ходе опыта число узлов (развилок) на одной ветви возрастало до 6 (рисунок). При этом количество информации (в битах), предположительно передаваемой разведчиками, было равно числу узлов дерева (поворотов) на пути к кормушке.
Лабиринт монтировался из спичек и укреплялся в кювете с водой для того, чтобы муравьи не могли двигаться по «прямой». В начальную точку муравьи попадали по мостику. Муравьи, находящиеся на арене, не могли видеть происходящего в лабиринте. Для того чтобы исключить маркировку пути пахучим следом, спички, по которым проходили муравьи, периодически заменяли новыми. В ходе эксперимента муравьев метили индивидуальными и групповыми метками. Опыты с разным числом развилок проходили в несколько сеансов. В каждом случае фиксировалось число правильных и ложных поворотов, совершаемых муравьями, а также измерялась общая длительность контактов разведчиков с муравьями.
§3. Определение скорости передачи информации
У большинства видов муравьев выделяются функциональные группы — разведчики и фуражиры. Разведчики патрулируют свои участки и, обнаружив пищу, приводят туда фуражиров из своей группы [ 13, 14 ].
В нашем эксперименте фуражировочная деятельность всех трех видов носила четко выраженный групповой характер. Муравьи-разведчики передавали информацию только внутри гнезда и всегда только постоянной группе из 5—8 фуражиров. Не все разведчики могли запомнить путь к кормушке, более того, число таких разведчиков уменьшалось в зависимости от сложности задачи: например, в случае двух развилок работало 5—6 разведчиков (и их групп) в течение сеанса, а в случае шести развилок — только 2. Для выяснения вопроса о передачи информации анализировали только те случаи, когда муравьи-фуражиры были лишены возможности подражать действиям разведчика: либо группа фуражиров двигалась далеко впереди него, либо сам разведчик был нами специально изолирован после контакта с фуражирами. Оказалось, что и по степени способности передавать информацию разведчики неоднородны. Группы, направляемые «бестолковыми» разведчиками, шли по неверному пути и, как правило, уходили в гнездо, не достигнув цели. С «перспективными» разведчиками и их группами мы работали специально в течение нескольких сеансов. Они не только сразу запоминали дорогу, но и сравнительно легко переучивались, когда в ходе одного сеанса менялось направление пути к приманке. Группы, контактировавшие с такими разведчиками, двигаясь компактно, безошибочно направлялись к приманке. Отдельные муравьи, совершавшие ложные повороты, довольно быстро догоняли остальных фуражиров. При этом члены других групп, в том числе «опытные» разведчики, попавшие на установку случайно и не знакомые с координатами кормушки в данный момент, не находили ее в случае трехи более развилок. Это позволяет исключить действие каких-либо иных механизмов ориентации, кроме дистантного наведения, т. е. использования сведений, сообщенных разведчиком.
Оценки коэффициентов уравнения линейной регрессии t = ai + b
и коэффициента корреляции r; х + Δ — 95%-ный доверительный
интервал для x
Вид |
a±Δ |
b±Δ |
r |
|---|---|---|---|
0,738 ±0,053 |
-0,763 ±0,094 |
0,962 |
|
| F. polyctena | 1,094 ±0,05 |
0,619 ±0,46 |
0,791 |
| C. saxatilis | 1,189 ±0,018 |
0,334 ±0,032 |
0,967 |
В нашем опыте с «бинарным деревом» количество информации (в битах) , необходимое для выбора правильного пути в лабиринте, равно числу развилок. Предположили, что время контакта разведчика с группой фуражиров (t) должно быть равно ai+b, где i — число развилок, а —коэффициент пропорциональности, равный скорости передачи информации (биг в минуту), а b — постоянная. Ее вводим потому, что муравьи могут передавать информацию, не имеющую прямого отношения к поставленной задаче, например сигнализировать «есть пища». По полученным данным оценивались параметры уравнения линейной регрессии а и b и вычислялся выборочный коэффициент корреляции (r) (см. табл. 1).
Оказалось, что у всех трех видов зависимость между временем контакта разведчика и фуражиров и количеством передаваемой информации (числом развилок i) близка к линейной, о чем свидетельствует большое значение выборочных коэффициентов корреляции (см. табл. 1). Это подтверждает нашу гипотезу (все коэффициенты корреляции достоверно отличаются от нуля при уровне значимости 0,01).
У вида ![]() F. sanguinea скорость передачи информации, т. е. величина а в уравнении
F. sanguinea скорость передачи информации, т. е. величина а в уравнении t=ai+b, равна 0,738 бит/мин, у F. polyctena — 1,094, у С. saxatilis — 1,189 бит /мин. Мы не считаем эти значения видовыми константами, вероятно, они могут варьировать. Заметим, что эти величины примерно на порядок меньше, чем скорость передачи информации у человека при разговоре [ 2 ].
§4. Теоретико-информационные тесты интеллекта муравьев
В описанной выше серии экспериментов использовалась случайная последовательность поворотов на пути к приманке — подбрасывали монету и выпадение «герба» означало поворот налево, «цифры» — направо. На основе того же метода «бинарного дерева» решили исследовать и другие вопросы, касающиеся языка и интеллекта муравьев. Одной из важнейших характеристик языка и интеллекта его носителей следует считать способность быстро подмечать закономерности и использовать их для кодирования, «сжатия» информации. Тогда длина сообщения о некотором объекте или явлении должна быть тем больше, чем оно сложнее. Например, человеку легче запомнить и передать такую последовательность поворотов на пути к цели: «ЛП ЛП ЛП ЛП ЛП» (здесь Л — налево, П —направо), чем более короткую, но случайную последовательность (допустим, «ПЛ ЛЛ ПП ЛП»).
Время, затрачиваемое муравьями-разведчиками  Formica sanguinea
Formica sanguinea
на передачу сведений о направлении к кормушке муравьями-фуражирами;
1-8 - «закономерные», 9-15 - случайные последовательности поворотов
№ пп |
Последовательность поворотов на пути к кормушке |
Среднее время, с |
Выборочное стандартное отклонение |
Число опытов |
|---|---|---|---|---|
1 |
ЛЛЛ |
72 |
8 |
18 |
2 |
ППП |
75 |
5 |
15 |
3 |
ЛЛЛЛЛ |
84 |
6 |
9 |
4 |
ППППП |
78 |
8 |
10 |
5 |
ЛЛЛЛЛЛ |
90 |
9 |
8 |
6 |
ПППППП |
88 |
9 |
5 |
7 |
ЛПЛПЛП |
130 |
11 |
4 |
8 |
ПЛПЛПЛ |
135 |
9 |
8 |
9 |
ЛЛП |
69 |
4 |
12 |
10 |
ЛПЛЛ |
100 |
11 |
10 |
11 |
ПЛЛЛП |
120 |
9 |
6 |
12 |
ППЛПЛ |
150 |
16 |
8 |
13 |
ПЛПППЛ |
180 |
20 |
6 |
14 |
ППЛППП |
220 |
15 |
7 |
15 |
ЛПЛЛПЛ |
200 |
18 |
5 |
Решили проверить, позволяет ли язык муравьев (и их интеллект) использовать простые закономерности «текста» для его «сжатия», где «текст» — последовательность поворотов на пути к кормушке. Результаты опытов приведены в табл. 2.
Так как не существует объективной количественной меры сложности текста (мера, предложенная в [ 12 ], алгоритмически невычислима), то, строго говоря, можем только проверить, совпадают ли представления о простом и сложном тексте у людей и муравьев. По-видимому, большинство людей сочтут, что последовательности поворотов в строках 5 и 6 в табл. 2 проще, чем в строках 7 и 8, а они в свою очередь проще, чем случайные последовательности той же длины (строки 13-15). Оказалось, что время передачи сведений о последовательности поворотов у муравьев также возрастает от простого к сложному (см. строки 5-8 и 13-15).
Статистическая значимость полученного результата может быть строго доказана. Сравним основную гипотезу H, согласно которой время передачи информации не зависит от сложности текста, с альтернативной гипотезой H1 о том, что время передачи тем больше, чем текст сложнее. Проведенный эксперимент включает 7 серий опытов с последовательностями поворотов одной длины (см. строки 5-8 и 13-15). Всего имеется 7! способов упорядочения этих семи серий; поэтому при справедливости гипотезы H опыт имеет 7! равновероятных исходов (упорядочивание выбирается в соответствии с убыванием времени передачи). Способы упорядочивания, при которых сперва идут самые простые последовательности поворотов 5 и 6, затем две промежуточные по сложности последовательности 7 и 8 и лишь после этого самые сложные последовательности 13-15, имеется 2! 2! 3!. Таким образом, при справедливости гипотезы H вероятность полученного в опыте упорядочения равна 2! 2! 3! / 7! = 1 / 210; малость этой вероятности заставляет отвергнуть гипотезу H и принять альтернативную гипотезу H1.
Интересно отметить, что муравьи начинают использовать закономерности для сжатия только достаточно больших текстов, например на передачу сведений о последовательностях длины «3» из 1- и 2-й строк табл. 2 тратилось даже чуть больше времени, чем на «случайную» последовательность той же длины из 9-й строки.
Литература
- Шеннон К. Математическая теория связи.— В кн.: Работы по теории информации и кибернетике. М.: Изд-во иностр. лит., 1963, с. 243—333.
- Яглом А. М., Яглом И. М. Вероятность и информация. М.: Наука, 1973.
- Линден 10. Обезьяны, человек и язык. М.: Мир, 1981.
- Панов Е. Я. Методологические проблемы в изучении коммуникации и социального поведения животных.— В кн.: Итоги науки и техники (зоология позвоночных). Вып. 12. М.: ВИНИТИ, 1983, с. 3-57.
- Меннинг О: Поведение животных. М.: Мир, 1982.
- Марков В. И., Островская В. М., Островский М. А. Структурный системный подход к анализу коммуникативных систем животных.—В кн.: Поведение животных в сообществах. М.: Наука, 1983, с. 4—6.
- Frisch К, von. The dance language and orientation of bees. Cambridge: Harvard University Press, 1967.
- Панов Е. Н. Эмпирический факт и его трактовка в этологии.— В кн.: Теоретические проблемы современной биологии. Пугцино, 1983, с. 104—119.
- Gardner R. A., Gardner В. Т. Teaching Sign Language to a Chimpanzee.— Science, 1969, v. 165, p. 664-672.
- Wassmann E. Die Psychischen Fahigkeiten der Ameisen.— Zoologica, 1899, H. 26, S. 1-133.
- Забелин С. И. Передача информации о направлении движения к приманке тактильным кодом у муравья Tapinoma erraticum.— Зоол. журн., 1979, т. 58, № 8, с. 1168-1172.
- Колмогоров А. Н. Три подхода к определению понятия «количество информации».— Пробл. передачи информ., 1965, т. 1, № 1, с. 3—11.
- Длусский Г. М. Принципы коммуникации у муравьев.— В кн.: Чтения памяти Холодковского. Л.: Наука, 1981, с. 3—33.
- Резникова Ж. И. Межвидовые отношения муравьев. Новосибирск: Наука. Сиб. отд-ние, 1983.
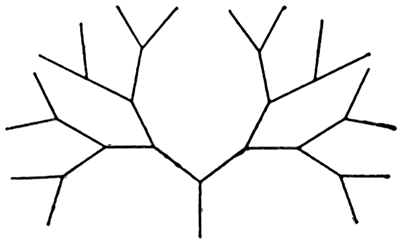
Схема лабиринта — «бинарного дерева» с четырьмя развилками
(от греч. ethos обычай, нравственный характер и ...логия), биологическая наука, изучающая поведение животных в естественных условиях; уделяет преимущественное внимание анализу генетически обусловленных (наследственных, инстинктивных) компонентов поведения, а также проблемам эволюции поведения.
(от лат. intellectus познание, понимание, рассудок), способность мышления, рационального познания. Латинский перевод древнегреческого понятия нус («ум»), тождественный ему по смыслу.
надсемейство летающих насекомых отряда перепончатокрылых, родственное осам и муравьям. Существует около 20 тысяч видов пчёл. Распространены на всех континентах, кроме Антарктиды. Пчёлы питаются нектаром и пыльцой, используя нектар главным образом в качестве источника энергии, а пыльцу для получения белков и других питательных веществ.
восприятие муравьями в качестве сигнала характерных движений других особей одной общины («язык поз»).
сигналы, передаваемые муравьями прикосновением антенн, щупиков, лапок при непосредственном контакте особей. Синоним: тактильный код.
 Formica sanguinea Latreille 1798 — кровавый муравей-рабовладелец
Formica sanguinea Latreille 1798 — кровавый муравей-рабовладелецмуравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
устойчивые группы особей, занятых в семье выполнением определенных функций. Входят в состав полиэтических групп.






