






Как было показано в предыдущем разделе, первым этапом формирования пустынной фауны было появление первичных упрощенных комплексов видов на базе, существовавшей в данной местности гумидной или семиаридной фауны. Такие первичные коадаптивные комплексы уже изначально имеют черты структуры, присущие любому пустынному комплексу, т. е. состоят из видов—герпетобионтов, отличающихся по питанию.
Как отмечалось выше, в гумидных областях спектры питания большинства видов муравьев сходны и возможность их сосуществования в одном биогеоценозе в первую очередь обеспечивается вертикальной (ярусной) дифференциацией. Поскольку в пустынях и сходных с ними азональных ландшафтах муравьи могут добывать пищу только в одном ярусе (на поверхности почвы), экологические ниши всех видов, входящих в первичный пустынный комплекс, в той или иной степени будут перекрываться. Этим в первую очередь, видимо, и объясняется бедность видового состава таких комплексов.
Из хорошо известной модели конкуренции между двумя видами Гаузе — Лотки — Вольтерра следует, что если факторы, определяющие численность двух видов, несколько различаются (т. е. если имеются различия в экологических нишах), возможно сосуществование этих видов. Эта модель не позволяет, однако, оценить, какая степень различий достаточна для такого сосуществования. Более того, из нее вытекает, что при любых самых малых различиях должна установиться равновесная двухвидовая система. Такое допущение, однако, противоречит всем известным фактам.
Первой и, насколько мне известно, единственной попыткой дать формализованное объяснение ограниченности видового разнообразия является модель Мэя и Мак-Артура (May, McArtur, 1972). Авторы рассматривают случай, когда в одном местообитании живет несколько видов, конкурирующих за какой-то ресурс, изменчивый по одному параметру, например, ряд зерноядных видов, питающихся семенами разного размера. Для каждого вида в этом ряду принимается некоторая функция использования ресурса (рис:90/а/), которая в данном конкретном примере определяется долей в питании семян определенного размера. Если форма этой функции описывается кривой нормального распределения и одинакова для разных видов, уровень межвидовой конкуренции можно задавать значением ω/d, где ω — среднее квадратическое отклонение функции использования ресурса, a d — расстояние между максимумами соответствующих функций. Этой идеальной картине довольно близко соответствуют описанные выше случаи дифференциации по питанию отличающихся размерами дневных зоонекрофагов (рис:3) и карпофагов (рис:6) пустынь.
При наличии видов с плотностями N1...Ni...Nn уравнениями конкуренции будут
где ki — соответствие среды потребностям 1-го вида (например, обилие на кормовом участке семян соответствующего размера), а αij — коэффициент перекрывания функций использования ресурсов i-го и j-го видов, лежащий в пределах от (пища совершенно различна) до 1 (функции использования ресурса полностью совпадают). Преобразовав это уравнение, мы получим
При t = ∞, т. е. в стабилизировавшемся многовидовом комплексе
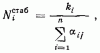
или, другими словами, в стабилизировавшемся комплексе видов, конкурирующих за ресурс, отличающийся одним параметром, плотность каждого вида будет прямо пропорциональна обилию ресурса с соответствующими параметрами и обратно пропорциональна давлению конкурирующих видов (степени перекрывания функции использования ресурса данного вида с функциями использования ресурсов других видов комплекса). Если принять все k и все α за независимые и постоянные величины, то в представленной модели, как и в модели Гаузе — Лотки — Вольтерра, система может стабилизироваться при любом числе видов. Картина, однако, меняется, если ввести, как это сделали Мэй и Мак-Артур, в модель вариабельность среды, приняв
где kj — постоянное среднее значение коэффициента, a γi(t) — гауссов «белый шум» с дисперсией σ². Для данного примера это означает случайную изменчивость в обилии семян разного размера. Оказалось, что при этом для устойчивости сообщества необходимо, чтобы ω/d было выше некоторого значения, зависящего от дисперсии «шума» и числа видов. Таким образом, согласно этой модели, число конкурирующих за общий ресурс, отличающийся по одному параметру, и обитающих совместно видов зависит от степени вариабельности среды. При этом чем сильнее меняется среда, тем больше должно быть расстояние между максимумами функций использования ресурсов совместно обитающих видов.
Если модель Мэя — Мак-Артура верна, мы должны ожидать, что вариабельность ресурсов в малопродуктивных экосистемах должна быть выше, чем в богатых и продуктивных. На самом же деле такая связь вовсе не является правилом. В качестве примера можно привести данные Н. Т. Нечаевой и К. Г. Антоновой (1970), которые в течение 8 лет (1960—1967 гг.) определяли ежегодную первичную продукцию в четырех растительных формациях в Центральных Каракумах (Каррыкуль). Как видно из данных табл.31, хотя средняя общая первичная продукция в тетыровой формации на такыровидных почвах была того же порядка, что и в формациях на песчаных почвах, колебания урожая по годам здесь были явственно больше. Особенно велики были здесь колебания урожая однолетних солянок (от 0,3 кг/га в 1961 г. до 313 кг/га в 1964 г.), злаков (от 0,3 кг/га в 1961 г. до 35,6 кг/га в 1966 г.) и эфемерового разнотравья (от 0,1 кг/га в 1961 и 1965 гг. до 14/4 кг/га в 1966 г.). В то же время колебания прироста Carex physodes, несмотря на очень низкую плотность, были относительно меньше, чем на участках с песчаной почвой. Точно так же не заметно явственной связи между средней продуктивностью растений и вариабельностью ее по годам в разных растительных формациях на песчаной почве. Сходная картина наблюдается и при сравнении вариабельности вторичной продукции. Так, в. 1970 и 1971 гг. В. А. Потоцкая (1975) проводила учеты Теперь рассмотрим, как согласуется приведенная модель с имеющимися фактическими данными. Полученные нами данные (табл.4, табл.5, табл.7, табл.10) указывают, что число совместно обитающих видов пустынных муравьев, различающихся размерами особей и питающихся пищей (семенами или трупами беспозвоночных), отличающейся главным образом по одному параметру (размерам), пропорционально богатству биоценоза. Еще ярче эта связь для зерноядных муравьев показана Дэвидсон (табл.11) и для зерноядных грызунов — Брауном (Brown, 1973). Иными словами, получается, что чем ниже количество доступной пищи (в конечном счете Σk), тем больше должно быть расстояние между максимумами функций использования ресурсов (d) и соответственно ниже уровень межвидовой конкуренции (ω/d) совместно обитающих видов в стабилизировавшемся коадаптивном комплексе. численности жуков в кронах саксаула на трех различных участках в Акыбае. Как видно из данных табл.32, наблюдается даже обратная корреляция — чем выше численность какой-либо группы жуков была на участке в 1970 г., тем больше различия между численностями в 1970 и 1971 гг. Вообще, если говорить в целом, то наибольшие колебания первичной продукции (как сезонные, так и по годам) в Средней Азии наблюдаются в глинистых эфемеровых пустынях. В то же время разнообразие муравьев-карпофагов здесь довольно велико, и уж во всяком случае выше, чем на солончаках, где отмечаются наиболее стабильные условия.
Несоответствие модели Мэя — Мак-Артура реально наблюдаемым фактам объясняется тем, что в ней делается ряд явно ошибочных допущений. Прежде всего подразумевается, что существование вида возможно при любой плотности. На самом же деле существует некоторый порог, при достижении которого вид начинает автоматически вымирать. Имеются по крайней мере три причины такого вымирания:
1) снижение размножаемости из-за малой вероятности встречи половых партнеров;
2) обеднение генофонда и снижение приспособленности популяции в целом;
3) случайное выпадение вида из-за случайных колебаний численности в результате неизбирательной элиминации при низкой плотности.
Априорно можно предполагать, что величины пороговой плотности будут разными у разных видов. Отсюда следует, что при плавном уменьшении коэффициента ki уравнения конкуренции (1) (т. е. фактически — продуктивности системы) должно уменьшаться и число совместно обитающих конкурирующих видов.
В модели Мэя — Мак-Артура коэффициенты (ki и αij) принимаются постоянными и независимыми. На самом деле они зависят как от плотности i-го вида, так и от плотности всех конкурирующих с ним видов ряда. Пределы использования ресурса — более или менее постоянное свойство вида. Так, например, для Messor оптимальный размер семян лежит где-то в пределах от зерна проса до зерна пшеницы. Более крупные семена ячменя или тыквы эти муравьи тащат уже с трудом. А слишком мелких семян, например мака, эти муравьи просто не замечают. Виды Monomorium, наоборот, собирают мелкие семена, с большим трудом доставляют в гнездо зерна проса и вообще не могут сдвинуть с места пшеничное зерно. Поэтому при изменении плотности вида меняется эксцессивность функции использования ресурса (рис:90/б/). Соответственно возрастает относительная доля ресурса со средними параметрами, т. е. меняется коэффициент ki. Больше того, при изобилии пищи муравьи (как и другие животные) будут предпочитать собирать пищу оптимального размера, что еще больше будет увеличивать эксцессивность функции использования ресурса. Как меняется коэффициент перекрывания ресурсов (αij) с изменением плотности одного из видов, хорошо видно на рис:90/б/. Через плотность коэффициенты ki и αij связаны между собой. Очевидно, в общем виде уравнение конкуренции может быть записано как
где Nimin — нижняя пороговая плотность вида, а Ki — коэффициент устойчивости, причем
где
Можно ожидать, что при некоторых условиях общее снижение количества доступного ресурса (Σk) должно приводить не только к постепенному выпадению из ряда видов, достигших пороговой плотности, но и к возникновению новой закономерной структуры ряда. Так, например, представим себе ряд видов А, В, С, D, Е, F, G с одинаковой плотностью и близкими, но неравными Nimin. Допустим, что происходит общее снижение плотности этих видов, и первым достигает пороговой плотности вид D. В результате исчезновения этого вида возрастает плотность видов С и Е (так как изменится Σα). В свою очередь, увеличение плотности этих видов вызовет увеличение коэффициентов αBC и αEF и соответственно снижение плотностей видов В и F. Хорошей аналогией этой ситуации является изменение активности видов в многовидовой ассоциации муравьев, построенной по иерархическому принципу, при изъятии одного из компонентов ассоциации. Так, в экспериментах Ж. И. Резниковой (1976) изоляция гнезд доминанта Formica pratensis привела к увеличению фуражировочной активности субдоминанта ![]() Formica cunicularia и одновременно к снижению активности инфлюэнтов Myrmica. При дальнейшем снижении плотности вероятность выпадения видов
Formica cunicularia и одновременно к снижению активности инфлюэнтов Myrmica. При дальнейшем снижении плотности вероятность выпадения видов B и F из ряда будет выше, чем видов С и Е. При других исходных условиях (разная исходная плотность, разная зависимость ki и αij от плотности, существенно различающиеся Nimin и т. д.) могут возникать и другие варианты структур. Однако обсуждение этих моделей за пределами как моей компетенции, так и задач, этой книги.
Из приведенных выше рассуждений следует вывод, имеющий прямое отношение к вопросу о путях адаптации. Вид может сохраниться в коадаптивном комплексе при увеличении конкурентного давления других видов (эволюционное совершенствование других членов комплекса, внедрение новых конкурентоспособных видов, общее снижение обеспеченности ресурсами и т. д.) только изменив один из двух независимых параметров — Nimin и K. Можно либо снизить нижнюю пороговую плотность вида (Nimin), либо увеличить коэффициент устойчивости (K).
Как отмечалось выше, причинами вымирания популяции при падении плотности ниже пороговой могут быть снижение размножаемости из-за малой вероятности встречи половых партнеров, обеднение генофонда и снижение приспособленности популяции в целом и случайное выпадение вида из-за случайных колебаний численности. Соответственно снижение нижней пороговой плотности может быть достигнуто следующими способами:
1. Повышение вероятности нахождения полового партнера.
Адаптации такого рода очень широко распространены у животных. К ним относятся совершенствование сигнализации, синхронизация вылета самок и самцов, по-видимому, достигающая крайней степени у многих пустынных муравьев, переход от равномерного к агрегированному распределению. У муравьев также наблюдается переход к спариванию внутри гнезда (Cardiocondyla, Alloformica) и размножение делением семей. Предел возможностей достигается у видов, переходящих к партеногенезу.
2. Выработка механизмов защиты от вредных последствий гомозиготности.
Как уже отмечалось выше, у муравьев, как и у других перепончатокрылых, наблюдается гаплодиплоидное определение пола, при котором вредные мутации элиминируются в первом же поколении. Можно предполагать, что такой способ определения пола возник именно как адаптация к существованию при низкой плотности популяций. Другой возможный путь защиты — увеличение плоидности, широко распространенный у растений.
3. Выработка адаптации, позволяющих снизить размах случайных колебаний численности.
Такие адаптации особенно хорошо заметны у социальных насекомых. Собственно само возникновение социальности и ее совершенствование в процессе эволюции являются процессами, ведущими к повышению гомеостатичности популяций. У пустынных муравьев мы наблюдаем также целый ряд специализированных приспособлений, повышающих гомеостатичность популяций в условиях пустынь: постройка глубоких гнезд, создание запасов пищи, эфемеризация брачного лёта.
Увеличение устойчивости может достигаться следующими путями:
1. Увеличение потенциально доступного ресурса (ki).
Собственно ресурсы способны увеличить лишь немногие виды, и одно из таких исключений — «разведение» муравьями тлей или грибов. Гораздо чаще увеличение достигается за счет изменений потенций вида. Для муравьев основной путь — это повышение эффективности использования кормового участка. Немаловажное значение для пустынных муравьев имеет, как было описано выше, увеличение вариабельности рабочей касты. У зерноядных муравьев увеличение ki может быть достигнуто за счет повышения эффективности усвоения корма, как было показано выше на примере Messor aralocaspius и M. denticulatus.
2. Уменьшение давления конкурирующих видов (Σαij).
Оно может быть достигнуто либо за счет перехода на питание ресурсом с другими параметрами (уменьшение размеров при отсутствии мелких конкурентов или увеличение — при отсутствии крупных), либо за счет специализации й сужения спектра питания. Второй путь должен обязательно сопровождаться одновременным повышением эффективности использования ресурса или снижением нижнего порога плотности. Основываясь на этих общих рассуждениях и данных о современной фауне, можно представить, как шла эволюция муравьев пустынь. Как было показано выше, вначале в азональных ландшафтах формировались первичные пустынные комплексы, состоявшие из видов, не имеющих специальных адаптации к жизни в аридных условиях. Межвидовая конкуренция на фоне нехватки пищевых ресурсов неизбежно должна была приводить к тому, что даже незначительные отличия в экологических нишах подхватывались и усиливались отбором. Виды, способные охотиться ночью, постепенно становились чисто ночными (Myrmecocystus, Camponotus). Виды, в питании которых семена составляли существенную долю, становились облигатными или преимущественными карпофагами (большинство![]() Myrmicinae аридных областей). Виды, не способные охотиться ночью или питаться семенами, оставались в прежней нише (дневные зоонекрофаги), но при этом приспосабливались добывать пищу в таких условиях, где большинство муравьев просто не смогло бы прокормиться. Одновременно должны были формироваться и специфические адаптации к жизни в пустыне, которые обеспечивали возможность сосуществования с конкурирующими видами в условиях с высокой температурой, низкой влажностью и резкими сезонными колебаниями количества пищи.
Myrmicinae аридных областей). Виды, не способные охотиться ночью или питаться семенами, оставались в прежней нише (дневные зоонекрофаги), но при этом приспосабливались добывать пищу в таких условиях, где большинство муравьев просто не смогло бы прокормиться. Одновременно должны были формироваться и специфические адаптации к жизни в пустыне, которые обеспечивали возможность сосуществования с конкурирующими видами в условиях с высокой температурой, низкой влажностью и резкими сезонными колебаниями количества пищи.
Положение усугублялось тем, что число видов, входящих в комплексы, все время увеличивалось. Действительно, в первичных аридных комплексах в одну трофическую группу, как правило, входит не более двух видов, имеющих разные размеры рабочих, тогда как в современных комплексах могут одновременно сосуществовать дневные зоонекрофаги пяти—шести и карпофаги трех—четырех различных размерных классов. Частично обогащение комплексов шло за счет симпатрического видообразования. Так, в пустынях Туранской низменности произошли виды группы Messor excursionis и подродов Monocombus и Eumonocombus рода Cataglyphis. Параллельно шел процесс миграции и взаимного обогащения комплексов, который хорошо прослеживается на примере Cataglyphis. Как показывают современные ареалы видов этого рода, существовало три или четыре вторичных очага видообразования, совпадающих с четырьмя основными аридными областями Палеарктики, существующими по крайней мере с миоцена. В пустынях Туранской низменности формировались подроды Monocombus и Eumonocombus, Иранского нагорья — группа C. albicans (подрод Cataglyphis s. str.), Северной Африки и Аравийского полуострова — подроды Machaeromyrma и Paraformica. Группа C. bicolor подрода Catagluphis s. str. скорее всего сформировалась в пустынях Индостана. Туранский, Северо-Африканский и Индийский аридные центры формообразования указываются и для рода Messor (Emery, 1912а; Bernard, 1973).
В настоящее время средние и крупные дневные зоонекрофаги (главным образом виды рода Cataglyphis) в большинстве биогеоценозов пустынь Палеарктики представлены смесью древних автохтонных элементов и потомков более поздних мигрантов, правда, зачастую прошедших автохтонную переработку. Так, например, в Центральной Сахаре (Delye, 1968) нишу ВЗ-3 занимает C. (Paraformica) emmae, ВЗ-4 — C. (Cataglyphis) albicans, ВЗ-5 — C. (Cataglyphis) bicolor, C. (Machaeromyrma) tucasi и C. (M.) bombicina. Первый и два последних вида являются прямыми потомками древнейшей западнопалеарктической (североафриканской или аравийской) пустынной фауны, C. albicans — вид иранского, а C. bicolor — индийского происхождения. Виды группы C. bicolor (C. bicolor и C. setipes) вообще широко распространены в настоящее время по всем пустыням Палеарктики. В пустынях Африки, Западной и Средней Азии они являются сравнительно недавними (возможно, плиоценовыми) пришельцами и теснят других крупных Cataglyphis — C. foreli в Средней Азии и Machaeromyrma — в Африке, Аравии и Западной Азии. За это время уже образовались два аллопатрических вида — C. bicolor и C. setipes, зона трансгрессии которых проходит через Закавказье и Копетдаг. В настоящее время активно идет дальнейшая дифференциация этих видов (Арнольди, 1964). Сравнительно недавними, плиоценовыми или даже послеледниковыми, вселенцами в пустыни Средней Азии являются также Pheidole pallidula, Monomorium kusnezovi, ![]() M. gracillimum, Crematogaster subdentata, Tapinoma karavaievi и Acantholepis semenovi. Все эти виды в Средней Азии не имеют близкородственных форм и относятся к группам, распространенным в Африке или Индии.
M. gracillimum, Crematogaster subdentata, Tapinoma karavaievi и Acantholepis semenovi. Все эти виды в Средней Азии не имеют близкородственных форм и относятся к группам, распространенным в Африке или Индии.
Постоянное усложнение комплексов неизбежно должно было сопровождаться увеличением специализации муравьев (уменьшением Σα). Однако специализация по типам питания (трупы насекомых, насекомые, активные ночью, семена растений) произошла уже на самых ранних этапах формирования комплексов. Поэтому главными путями специализации могли быть временная (в пустынях с двумя максимумами осадков), пространственная (адаптации к жизни в разных типах пустынь) и в первую очередь размерная дифференциация. При отсутствии крупных видов с каким-либо типом питания видообразование должно сопровождаться увеличением размеров рабочих части видов, а при отсутствии мелких — наоборот. Неслучайно поэтому, что в Южной Америке самые крупные виды Pheidole встречаются именно в пустынях (Kusnezov, 1956). Довольно крупные размеры Anoplolepis пустынь Южной Африки явно вторичны. У этих муравьев антенны не 12-, а 11-члениковые, а, как известно, редукция числа члеников антенн происходит у муравьев при уменьшении абсолютных размеров рабочих (Bunzli, 1935). Косвенно это подтверждается и тем, что Anoplolepis сицилийского янтаря (миоцен) были очень мелкими (Emery, 1891). Явно вторичны и крупные размеры Chelaner австралийских пустынь, поскольку подавляющее большинство представителей рода Monomorium (предки Chelaner) отличается мелкими размерами и исключение составляют лишь некоторые специализированные виды, обитающие в аридных и семиаридных областях Азии и Африки.
Выход в свободную трофическую нишу может быть осуществлен без особых морфофизиологических преобразований. Для этого достаточно изменить средние размеры рабочих (размерная дифференциация), температурный оптимум фуражировки (сезонная временная дифференциация). Сложнее осуществляется переход в другую стацию. Так, специализированные обитатели песчаных пустынь часто обладают сложным псаммофором, приспособлениями для отбрасывания песка на передних ногах, светлой окраской, у них встречаются наиболее сложные гнезда и т. д. Однако наиболее сильных перестроек требует внедрение в уже существующий комплекс. В этом случае уменьшение давления конкурирующих видов (Σαij) может быть достигнуто только за счет сужения спектра питания. Но при этом, как уже отмечалоь выше, одновременно должна повышаться эффективность использования ресурса (увеличение ki) и (или) снижаться нижний порог плотности вида (Nimin). Поэтому на практике мы значительно чаще наблюдаем, что вновь вселяющиеся виды, внедряясь в уже существующий комплекс, вытесняют один из видов уже сложившегося ряда. Как правило, это происходит в том случае, если вселяющийся вид обладает более высокой социальной организацией и, следовательно, меньшей пороговой плотностью (за счет большей гомеостатичности семей) и большей эффективностью использования кормового ресурса (за счет более мобильной и эффективной системы использования кормового участка). Вытесненный вид сохраняется лишь в стациях, где он в силу каких-то особенностей поведения оказывается более конкурентоспособным. Иллюстрацией к этому могут быть описанные выше взаимоотношения между Proformica epinotalis и Acantholepis semenovi или Cataglyphis foreli и C. setipes.
Все рассмотренные выше адаптации муравьев к жизни в пустыне по сути дела являются адаптациями, возникающими в результате межвидовой конкуренции. По всей видимости, обнаруженная закономерность вообще характерна для эволюции всех живых организмов. Неслучайно архаичные реликтовые виды—это, как правило, виды, не имеющие конкурентов (т. е. «выпавшие» из коадаптивных комплексов). Отсутствие конкурентов может объясняться либо географической изоляцией, либо необычной специализацией. Характерный пример консервирующего действия изоляции — гаттерия, единственное наземное позвоночное мелких островов у побережья Новой Зеландии. Примеры сохранения реликтовых муравьев Alloformica aberrans и Aphaenogaster raphidiiceps в результате специализации описаны выше. Еще более яркие примеры можно найти в других группах животных. Так, пилильщики из рода Xyela, представители наиболее древнего семейства перепончатокрылых, известного с верхнего триаса, обитают в соцветиях хвойных, где почти не встречаются другие насекомые. Двоякодышащие рыбы, известные со среднего девона, в настоящее время представлены десятком аллопатрических видов, причем все они обитают в периодически пересыхающих водоемах, бедных кислородом. Щитни (Notostraca), известные с триаса (современный вид Triops cancriformis), обитают во временных водоемах. Необычайно высокая скорость роста и способность расселяться с помощью ветра на большие расстояния (разносятся очень мелкие покоящиеся яйца) позволяют этим довольно крупным ракообразным заселять водоемы, где у них нет ни врагов, ни конкурентов.
Все рассмотренные выше примеры показывают также, что возникновение новых видов и родов (т. е. макроэволюционные преобразования) могло происходить только в условиях коадаптивной эволюции. В то же время хорошо известно, что многие микроэволюционные преобразования могут происходить и при отсутствии межвидовой конкуренции. Примером тому могут служить хотя бы хорошо изученные случаи выработки устойчивости к ядам у насекомых, или возникновение индустриального меланизма. Видимо, можно принять, что именно в этом заключается различие в механизмах микро- и макроэволюции. В таком случае если элементарной единицей микроэволюции является популяция, то элементарной единицей макроэволюции — коадаптивный комплекс.
Имеется и другое очень существенное различие между микро- и макроэволюцией. Микроэволюционные преобразования часто бывают случайными и ненаправленными. Это связано с тем, что на таком уровне необходимый результат может быть достигнут различными путями. Конкретный результат будет определяться генофондом популяции, в которой идет отбор. Напротив, макроэволюция, как правило, направлена. Это хорошо видно на примере коадаптивных комплексов муравьев пустынь Земли. Независимо и параллельно возникали комплексы, удивительно сходные по структуре, причем виды, занимающие сходные ниши в этой структуре, обладают конвергентным морфологическим сходством. Оно может быть настолько велико, что еще недавно объединялись роды, относящиеся к разным трибам. На нашем материале хорошо видны и причины такой направленности. Ими являются исходное сходство различных муравьев-герпетобионтов в гумидных и семиаридных областях Земли, сходство условий в различных пустынях, определяющее жесткие рамки возможной структуры комплексов муравьев, и общие закономерности возникновения и эволюции коадаптивных комплексов.
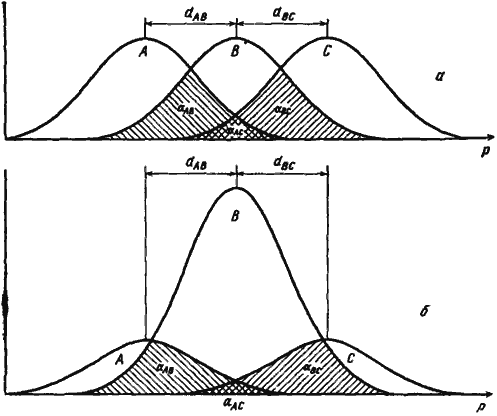
Рис. 90. Функции использования ресурса группой видов (А, В, С), ресурсы которых отличаются по одному параметру (Р) и расстояния между максимумами функций которых (d) равны
а — при равной численности;
б — при различной численности.
Объяснения в тексте
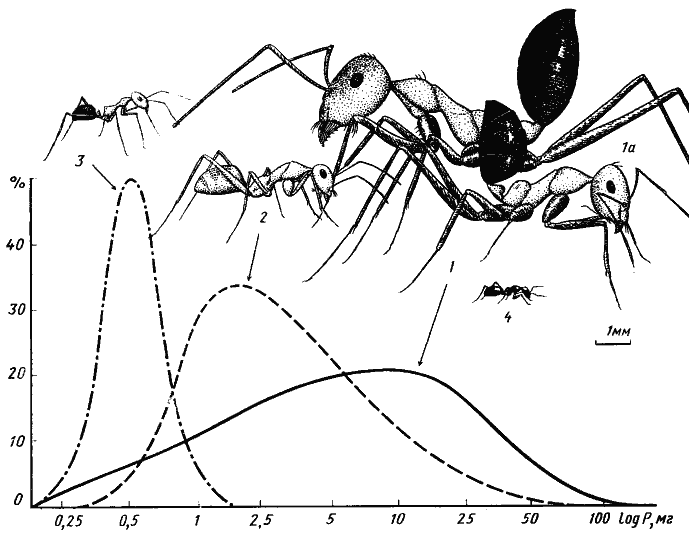
Рис. 3. Сравнительные размеры четырех основных видов дневных зоонекрофагов песчаной пустыни (Акыбай) и частотное распределение (в %) добычи трех видов по весовым классам (усредненные данные)
1, 1а — средний и крупный рабочие Cataglyphis setipes;
2 — C. pallida;
3 — Acantholepis semenovi;
4 — Plagiolepis pallescens
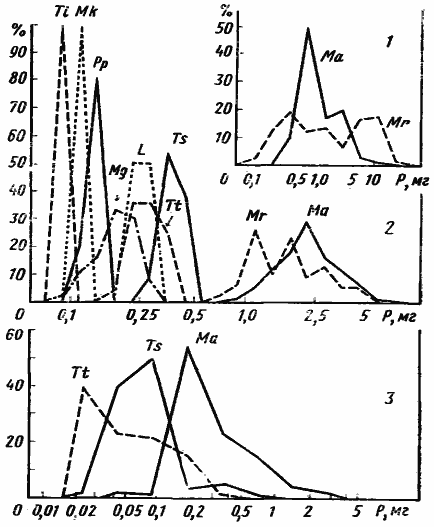
Рис. 6. Дифференциация по размерам и питанию у муравьев-карпофагов, совместно обитающих в подгорной степи
(Варганза)
1 — частотное распределение по весовым классам грузов отдельных фуражиров Messor aralocaspius (сплошная линия) и М. rufitarsis (пунктир); 2 — частотное распределение фуражиров разных видов муравьев по весовым классам; 3 — частотное распределение по весовым классам семян в питании трех массовых видов. Размеры грузов и размеры семян в питании Messor не совпадают, так как эти муравьи часто собирают плоды или колоски с многими семенами. Шкала логарифмическая; размеры весовых классов: lg 2 (1, 3), lg 1,26 (2) L — Leptothorax semenovi; Ma — Messor aralocaspius; Mr — M. rufitarsis; Mk — Monomorium kusnezovi; Mg —
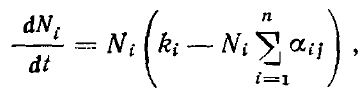
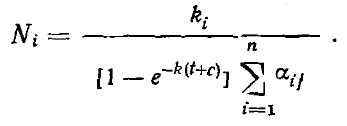
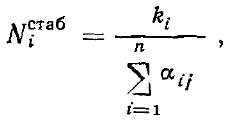
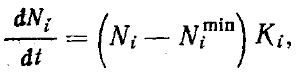
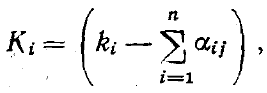
организм, обитающий среди растительных или иных органических остатков, на поверхности почвы.
эволюционно сложившаяся, пространственно ограниченная, длительно самоподдерживающаяся, однородная экологическая система, в которой функционально взаимосвязаны живые организмы и окружающая их абиотическая среда.
(от греч. oikos дом, жилище, местопребывание и ...логия), наука об отношениях живых организмов и образуемых ими сообществ между собой и с окружающей средой. Термин «экология» предложен в 1866 Э. Геккелем. Объектами экологии могут быть популяции организмов, виды, сообщества, экосистемы и биосфера в целом.
(зоонекрофаг), организм, питающийся мертвыми животными, гетеротроф
организм, питающийся семенами растений.
участок, используемый муравьями для добычи пищи.
животные, не имеющие позвоночника: простейшие, губки, кишечнополостные, черви, моллюски, членистоногие, иглокожие и др.; примерно 3/4 видов всех животных.
совокупность особей одного вида, длительно занимающая определенное пространство и воспроизводящая себя в течение большого числа поколений. В современной биологии популяция рассматривается как элементарная единица процесса эволюции, способная реагировать на изменения среды перестройкой своего генофонда.
Всего 106 видов.
территориальная иерархическая группировка муравейников разных видов, обладающая устойчивой структурой, которая поддерживается поведенческими механизмами.
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
 Formica cunicularia Latreille, 1798 — прыткий степной муравей
Formica cunicularia Latreille, 1798 — прыткий степной муравейнеобратимое историческое развитие живой природы. Определяется изменчивостью, наследственностью и естественным отбором организмов. Сопровождается приспособлением их к условиям существования, образованием и вымиранием видов, преобразованием биогеоценозов и биосферы в целом.
Всего 38 видов.
Всего 4 вида.
обязательное существование только сообществами (семьями, общинами).
подотряд насекомых отряда равнокрылых. Длина 0,5-6 мм. Св. 2,5 тыс. видов. Распространены главным образом в умеренном поясе Северного полушария. Живут колониями на растениях. Многие тлевые опасные вредители, некоторые переносчики ряда вирусных заболеваний растений. Сахаристые испражнения тлевых (падь) привлекают муравьев, которые охраняют тлевых.
у муравьев имеется три основные касты: самцы, самки и рабочие (бесплодные модифицированные самки).
ксеробионт
гигрофил
Всего 30 видов.
Всего 965 видов.
итал. obligato, "обязательный", необходимый, существенный
Myrmicinae — самое большое подсемейство муравьев, включающее более 2 тыс. видов, относящихся более чем к 150 родам.
Всего 65 видов.
ксеробионт
ксеробионт
Старое название: Pheidole arenarum Ruzsky
галобионт
Subspecies: Monomorium indicum kusnezowi Santschi, 1928
 Monomorium destructor (Jerdon, 1851) — изящный муравей, муравей-разрушитель
Monomorium destructor (Jerdon, 1851) — изящный муравей, муравей-разрушительСтарое название: Monomorium gracillimum
гигрофил, галобионт
Старые названия: Acantholepis semenovi, Acantholepis transcaspiensis
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
Всего насчитывает 549 видов.
«медовые муравьи»
Всего 22 вида.
специальное приспособление для переноски сухой почвы, распространенн у пустынных муравьев, образован длинными отстоящими волосками (аммохетами), часто имеющими форму латинской буквы J, расположен на нижней поверхности головы.
олигогинны
Старое название: Leptothorax semenovi
Старое название: Messor rufitarsis
Старые названия: T. chefteki, T. turcomanica, T. turcmenicum, T. taurocaucasum, T. taurocaucasicum






