






Трофобиоз муравьев с различными видами Homoptera является одной из форм симбиоза животных. Впервые он был детально изучен А. К. Мордвилко (1901). В последние годы, в связи с решением ряда практических вопросов, было получено много новых интересных сведений об этом явлении. Опубликован ряд работ обзорного характера (Nixon, 1951; Wellenstein, 1951, 1952; Гринфельд, 1961).
Значение трофобиоза для муравьев. Ряд видов равнокрылых хоботных (Homoptera), преимущественно тлей (Aphidinea), а также кокцид (Coccinea), листоблошек (Psyllinea) и цикадок (Cicadinea), выделяет большое количество экскрементов с высоким содержанием углеводов. Так, в сухом веществе экскрементов тлей содержится 85% углеводов и 3% (или 1—1,3% к общему весу) белка (Кузнецов, 1948; Кауцис, 1956). Состав компонентов у разных видов различен. Так, экскременты Cinara peclinatae Nordl. состоят в основном из глюкозы и небольшого количества сахарозы, а Stomaphis quercus L. — преиму-щественно из сахарозы (Gontarsky, 1940) /Здесь, как и всюду в настоящей работе, названия тлей даются согласно синонимике Г. Х. Шапошникова в «Определителе насекомых Европейской части СССР», т. 1. М.— Л., стр. 489—616/.

Эти экскременты, на-зываемые падью или медвяной росой, являются одним из основных компонентов пищи большинства видов му-равьев (рис:87). В на-стоящее время в Европе зарегистрировано 39 видов хоботных (35 — тли, 3 — червецы, 1 — листоблошки), с которыми связаны виды рода Formica (табл.15). Представители подрода Formica s. str. связаны главным образом с на-иболее продуктивными видами тлей из семейства Lachnidae, живущих на деревьях, преимущественно на хвойных. Представители других подродов связаны больше с тлями, живущими на травянистых растениях. Исключение составляют F. cinerea и F. exsecta, которые в лесах всегда предпочитают Lachnidae, но и эти виды чаще, чем Formica s. str., используют тлей с травянистых растений.
ТАБЛИЦА 15
Равнокрылые хоботные, с которыми связаны муравьи из рода Formica
(Западная Европа, Европейская часть СССР)
|
Вид растения
|
Хоботные
Вид (подотряд, семейство) |
Подроды муравьев
|
Автор
|
| Сосна обыкновенная | Cinara pini L. (Aph., Lach.)* |
F
|
Zoebelein, 1956; Muller, 1960 |
|
»
|
С. nuda Mordv. (Aph., Lach.)* |
F
|
Donisthorpe, 1915; Zoebelein, 1956; Ehrhardt, etc., 1961 |
|
»
|
C. pinihabitans Mordv. (Aph., Lach.)* |
F, S, C
|
|
| Сосна горная | C. neubergi Arnh. (Aph., Lach.) |
F
|
Zoebelein, 1956 |
| Кедр сибирский | C. cembrae Choi. (Aph., Lach.) |
F
|
Zoebelein, 1956 |
| Ель | C. grossa Kalt. (Aph. Lach.)* |
F, R
|
Gontarsky, 1940; Zoebelein, 1956; Ehrhardt, etc., 1961; Scheurer 1965 |
|
»
|
C. pilicornis Hart. (Aph., Lach.)* |
F, S
|
Ehrhardt etc., 1961; Scheurer, 1965 |
|
»
|
C. cistata Buckt. (Aph., Lach.) |
F
|
Zoebelein, 1956 |
|
»
|
C. costata Zett. (Aph., Lach.)* |
F, S
|
Scheurer, 1965 |
|
»
|
C. bogdanovi Mordv. (Aph., Lach.) |
F, R, S
|
Scheurer, 1965 |
|
»
|
Physokermes piceae Schrk. (Coc, Coccid.) |
F, R, S
|
Zoebelein, 1956 |
| Лиственница европейская |
C. laricis Walk. (Aph., Lach.) |
F
|
Gontarsky, 1940 |
| Пихта | C. pectinatae Nordl.(Aph.,Lach.) |
F
|
Gontarsky, 1940 |
| Бук | Lachnus pallipes Hart. (Aph., Lach.) |
F
|
Schouteden, 1902; Zoebelein, 1956; Muller, 1960; Schmutterer, 1956 |
| Дуб | L. roboris L. (Aph., Lach.) |
F, S
|
Zoebelein, 1956; Schmutterer, 1958 |
|
»
|
Stomaphis quercus L. (Aph., Lach.) |
F
|
Gontarsky, 1940 |
| Береза | Symydobius oblongus Heyd. (Aph., Call.)* |
F, S
|
|
| Осина | Chaitophorus populeti Panz. (Aph., Chait.)* |
F, S, C
|
|
|
»
|
Ch. albus Mordv. (Aph., Chait.)* |
F
|
|
| Ольха | Pterocallis maculata Heyd. (Aph., Call.)* |
C
|
|
| Калина | Aphis viburni Scop. (Aph., Aphid.)* |
C
|
|
| Рябина | Disaphis sorbi Kalt. (Aph., Aphid.)* |
F
|
|
| Боярышник | Psylla crataegi Schrnk (Ps., Psyll.) |
F
|
Zoebelein, 1956 |
| Различные яблоневые (яблоня, боярышник, рябина и др.) |
Aphis pomi Deg. (Aph., Aphid.)* |
F, C
|
Zoebelein, 1956 |
| Мать-и-мачеха | Anuraphis farfarae Koch. (Aph., Aphid.) |
S
|
Schouteden, 1902 |
| Подорожник | Aphis plantaginis Goetze (Aph., Aphid.) |
F
|
Donisthorpe, 1915 |
| Вереск | Arctorthezia cataphracta Shaw. (Coc, Orth.) |
S
|
Donisthorpe, 1915 |
| Конский щавель | Aphis fabae Scop. (Aph., Aphid.)* |
C, R
|
|
| Лютики | Dysaphis ranunculi Kalt. (Aph., Aphid.) |
S
|
Donisthorpe, 1915 |
| Сложноцветные | Trama radicis (Schrk.) (Aph., Aphid.) |
S
|
Donisthorpe, 1915 |
|
»
|
Aphis sp. (Aph., Aphid.)* |
S
|
|
| Корни злаков | Forda formicaria Heyd. (Aph., Lach.) |
S
|
Donisthorpe, 1915 |
|
»
|
F. marginata Koch.(Aph., Lach.) |
S
|
Donisthorpe, 1915 |
|
»
|
F. trivialis Pass. (Aph., Lach.) |
R
|
Donisthorpe, 1915 |
|
»
|
Parachletus cimiciformis (Aph., Lach.) |
F
|
Gosswald, 1934—1935 |
|
»
|
Tetraneura ulmi L. (Aph.,Lach.) |
S
|
Donisthorpe, 1915 |
|
»
|
Anoecia corni F. (Aph., Anoec.) |
F, S
|
Donisthorpe, 1915 |
| Различные растения | Newstedia floccosa Dog. (Coc, Orth.) |
F, S
|
Donisthorpe, 1915 |
| Клен | ? (Aph.)* |
F
|
Примечание. Названия тлей даны в соответствии с синонимикой Г. X. Шапошникова. Все данные о связи муравьев рода Formica с хоботными на территории СССР (отмечены звездочками) собраны автором в Московской и Владимирской областях в 1956—1965 гг. и публикуются впервые. Сокращения: Anoec. — Anoeciidae, Aph. — Aphidinea, Aphid. — Aphididae, C — Coptoformica, Call. — Callaphidiidae, Chait. — Chaitophoridae, Coc. — Coccinea, Coccid. — Coccidae, F — Formica s. str., Lach.— Lachnidae, Orth. — Ortheziidae, Ps. — Psyllinea, Psyll. — Psyllidae, R — ![]() Formica sanguinea, S — Serviformica.
Formica sanguinea, S — Serviformica.
Одна семья рыжих лесных муравьев из 100 тыс. особей в течение 100 дней собирает около 10 кг пади, причем за день с одного дерева муравьи приносят в среднем около 60 г (Oekland, 1930 a,b, 1932). Более детальные исследования Цеблайна (Zoebelein, 1954, 1956) показали, что разные ви¬ды собирают различное количество пади. Семья ![]() F. rufa собирает за год в среднем 450—500 кг пади (или 90—100 кг сахара), а F. polyctena — 290—300 кг пади (60—65 кг сахара).
F. rufa собирает за год в среднем 450—500 кг пади (или 90—100 кг сахара), а F. polyctena — 290—300 кг пади (60—65 кг сахара).
Продуктивность различных видов тлей не одинакова. Так, за лето с колоний Cinara nuda Mordv. на соснах рыжие лесные муравьи собирают 70 кг пади с дерева за лето, с колоний Cinara piceae Panz. на ели — 30 кг с дерева, с колоний Lachnus (Schizodryobius) pallipes Hart, на буке 25— 30 кг с дерева (Zoebelein, 1956). В течение сезона продуктивность разных видов также меняется. В лесах на юге ФРГ весной F. polyctena собирают падь, главным образом, на ели (C. piceae), летом — на лиственнице (C. laricis Walk.), а во второй половине лета и осенью — на буке (L. pallipes) (Н. Muller, 1960).
Значение трофобиоза для тлей. Уже давно было замечено, что колонии тлей, на которых муравьи собирают падь, находятся в гораздо лучшем со¬стоянии, чем колонии тех же видов, не посещаемые муравьями. Даже про¬стой учет тлей показывает, что численность Lachnidae в лесах в несколь¬ко раз увеличивается вблизи муравейников рыжих лесных муравьев (Кауцис, 1956; Schmutterer, 1956; Н. Muller, 1958).
Сведения о том. что муравьи активно уничтожают естественных врагов тлей (Мордвилко, 1901 и др.), как показывают современные данные, значительно преувеличены (Nixon, 1951). Более или менее эффективно Formica защищает тлей от наездников, гораздо меньше от личинок Syrphidae и совсем не защищают от божьих коровок (Wellenstein, 1952). Однако А. К. Мордвилко (1901) отмечал, что муравьи не предохраняют тлей от наездников. По нашим наблюдениям, муравьи не уничтожают личинок Leucopis (Diptera).
Кроме того, основным врагом тлей, например L. pallipes, часто являются птицы, поедающие зимующие яйца (Schmutterer, 1956).
Деятельность муравьев вызывает увеличение продуктивности тлей и скорости их развития (El-Ziady, Kennedy, 1956; El-Ziady, 1960; Banks, 1958). Однако различий в колониях тлей, посещаемых и не посещаемых муравьями, живущих на листьях одного возраста не обнаружено. По-видимому, разница в скорости развития и продуктивности вызывается тем, что муравьи переносят тлей на молодые части растений.
Известно также, что муравьи не только переносят тлей на более подходящие части растений, но и строят над ними галереи, защищающие их от врагов и прямых солнечных лучей (Мордвилко, 1901, 1936, и др.), и уносят зимующие яйца в глубь гнезда (Zwolfer, по Гринфельду, 1961). Мы наблюдали самок тлей в центре зимующего «клуба» из рабочих ![]() F. fusca.
F. fusca.
Далеко не все виды тлей находятся в симбиозе с муравьями. Тлей, покрытых восковым или лаковым налетом, или живущих в галлах, муравьи не охраняют. Как показали исследования Э. К. Гринфельда (1961), в экскрементах этих тлей сахара отсутствуют. Тли, выделяющие падь и охраняемые муравьями, обладают рядом морфологических приспособлений к симбиозу. Так, например, у них сильно развиты перианальные волоски, на которых удерживается капля экскрементов, редуцированы играющие защитную функцию соковые трубочки и «хвостики» (Мордвилко, 1936; Н. Schmidt, 1952; Kloft, 1959а).
Присутствие муравьев, как показали опыты с применением радиоактивного фосфора, вызывает усиление экскреции у тлей почти вдвое (Banks, Nixon, 1958). Растения Vicia fabae выращивались в водной культуре с солями, содержащими P³². Радиоактивность тлей, которых посещали муравьи, была вдвое выше, чем контрольных. После выделения капли экскрементов радиоактивность падала и возвращалась к прежнему уровню приблизительно через 20 мин. Когда эти колонии изолировали от муравьев, экскреция их постепенно падала до уровня контрольных колоний. Высасывание соков растений у тлей происходит при помощи своеобразного «насоса», помещенного в голове. В нормальных условиях этот «насос» работает с определенными интервалами, и при наличии муравьев эти интервалы укорачиваются.
Причины высокого содержания сахаров в экскрементах тлей до сих пор не ясны. Наиболее распространенной является гипотеза Бюсгена (по Гринфельду, 1961 и Гилярову, 1948), согласно которой тли «вынуждены» пропускать большое количество сока растений, для того чтобы получить необходимое количество белка. Сахара же остаются в избытке и выделяются с экскрементами. Однако имеется ряд фактов, не согласующихся с этой гипотезой.
1. Экскременты тлей содержат до 3% белка, а в соке растений его мало, всего 5% (по Кузнецову, 1948).
2. Нет сахаров в экскрементах сосущих насекомых, не связанных с муравьями, хотя и живущих на тех же растениях. Возможно, они идут на образование воска или лака, но их нет и у тлей, живущих в галлах и не выделяющих защитных покровов (Гринфельд, 1961).
3. У тлей имеются симбионты, способные, по-видимому, фиксировать азот воздуха (Штейнгауз, 1952).
М. С. Гиляров (1948) выдвинул другую гипотезу, согласно которой тли, не покрытые защитным восковым или лаковым слоем, «вынуждены» для поддержания нормального водного баланса поглощать большое количество сока. Большая часть воды при этом испаряется, а сахара и другие вещества выделяются с экскрементами. Эта гипотеза подтверждается опытами этого автора с Aphis fabae Scop.
Согласно третьей гипотезе, выдвинутой Э. К. Гринфельдом (1961), выделение сладких экскрементов является приспособлением тлей к симбиозу с муравьями, выработанным в результате длительной совместной эволюции, подобно железам многих мирмекофилов (Lycaenidae, Pselaphidae, некоторые Staphylinidae), выделяющих различные вещества, употребляемые муравьями в пищу.
Скорее всего правомочны обе гипотезы, однако, несомненно, нужно признать приспособление к защите от высыхания первичным.
Практическое значение связи муравьев с тлями. Как известно, многие тли являются серьезными вредителями растений. Поэтому при оценке экономического значения муравьев необходимо оценить их влияние на растения через посредство тлей, которых они охраняют.
Как показали исследования Клофта (Kloft, 1960с), тлей, живущих на деревьях, можно разделить на две группы: тли, сосущие из паренхимы, и тли, сосущие из флоэмы. Представители первой группы производят падь в небольшом количестве и поэтому не используются рыжими лесными муравьями. Точно так же муравьи не связаны с тлями, покрытыми восковым налетом. В то же время к этим группам и относятся почти все основные тли, вредящие лесу. Рыжие лесные муравьи в лесах используют выделения тлей, сосущих из флоэмы. Сюда относятся и представители се-мейства Lachnidae, предпочитаемые муравьями в хвойных лесах. Вред же от этих тлей, особенно по сравнению с видами, сосущими из паренхимы, ничтожен (Gosswald, 1941а; Kloft, 1953). Но при оценке экологического значения какой-либо группы нельзя принимать во внимание только один фактор. Как известно, основной вред от тлей на деревьях часто вызывается не тем, что они сосут сок, а тем, что своими экскрементами залепляют устьица листьев и вызывают распространение сажистого грибка. Муравьи же, собирая экскременты, предотвращают эту опасность (по Халифману, 19636). Опыты А. Р. Кауциса (1956) показывают, что стекла, положенные под деревьями, не посещаемыми муравьями, покрываются капельками экскрементов, а при наличии муравьев остаются чистыми.
Кроме того, следует учитывать, что муравьи в первую очередь охотятся именно на тех деревьях, которые они посещают из-за тлей (см. выше).
Немаловажное значение имеет падь как источник дополнительного питания для хищных и паразитических насекомых — наездников, сирфид и др. У этих насекомых увеличивается продолжительность жизни и плодовитость. Всего в ФРГ зарегистрировано около 300 видов насекомых, питающихся падью, 2/3 из которых полезны для леса (Zoebelein, 1955, 1957; Wellenstein, 1957с).
Основным критерием оценки роли муравьев являются изменения в состоянии деревьев, на которых они разводят тлей. Даже многолетнее разведение Lachnidae на одних и тех же елях, находящихся дальше 11 м от гнезд рыжих лесных муравьев, не влияет на состояние деревьев; лишь несколько елей, находящихся ближе 10 м от гнезда, снижают годовой прирост на 10% (Wellenstein, Muller, 1954). На прирост сосен разведение тлей вообще не оказывает влияния (Кауцис, 1956).
Однако даже Lachnidae, по-видимому, могут причинять вред молодым лесам и вызывать деформацию побегов, а в ряде случаев и гибель молодых елок (Н. Muller, 1956а, 1960, и др.).
По наблюдениям, проведенным в ГДР и ФРГ, разводимая на дубах рыжими лесными муравьями тля Lachnus roboris L. не причиняет вреда деревьям, но в Венгрии отмечены случаи, когда эта тля служила переносчиком язвы дубов (Schmutterer, 1956). По нашим наблюдениям, в Московской и Воронежской областях муравьи крайне редко разводят тлей на дубах, а чаще кормятся там выделяющимся из трещин соком. Единственную, действительно серьезную опасность в лесах представляют буковые тли (Lachnus (Schizodryobius) pallipes (=L. longirostris, L. exciccator), охраняемые рыжими лесными муравьями. Эти тли вызывают дегенерацию камбия и образование трещин коры у буков. В свою очередь это ведет к заражению деревьев буковым язвенным грибком Nectria ditissima: на старых деревьях в результате отмирают крупные ветви, а молодые часто погибают. Поэтому в буковых и смешанных (хвойные и бук) лесах рыжие лесные муравьи приносят серьезный вред (Н. Muller, 1956а, 1958; Schmutterer, 1956; Wellenstein, 1957с).
В садах тли также являются серьезными вредителями. Рыжие лесные муравьи, охраняя этих тлей, в частности яблоневую тлю Aphis pomi Deg., могут приносить в садах большой вред и являются здесь, безусловно, вредными насекомыми.
Но не следует забывать и о том, что падь тлей служит пищей для имаго многих хищных и паразитических насекомых, главным образом перепончатокрылых (наездников, ос) и двукрылых. В настоящее время насчитывается около 300 видов насекомых, питающихся падью, из которых 2/3 являются полезными (Zoebelein, 1956; Wellenstein, 1957b).
В последние годы в ФРГ все большее внимание уделяется лесному пчеловодству. В связи с тем, что луга с медоносами оказались распаханными, пасеки переносят в леса. В этих условиях пчелы собирают большое количество падевого меда. Наши пчеловоды отрицательно относятся к падевому меду, поскольку в ряде случаев наблюдается отравление пчел падью. Но ядовита лишь падь с некоторых растений (дуба, бархатного дерева, амурской сирени, ив). Мед же, полученный из пади лахнид, живущих на хвойных (Cinara sp. sp.), немногим уступает по качеству цветочному (Wellenstein, 1957b, 1958, 1960; Wellenstein, Muller, Kaeser, 1957; Kloft, 1960b).
Вблизи крупных колоний рыжих лесных муравьев (особенно F. polyctena) урожай падевого меда заметно увеличивается. На юго-западе ФРГ с середины июня до конца августа в 1958 и 1959 гг. проводилось обследование 40 пасек. Оказалось, что в лесах без муравьев среднее количество взятка на улей в 1,6 раз больше, чем на полях, а в лесах с муравьями — в 2,4 раза. Во всех случаях пчелы из пасек, расположенных у края леса, давали в среднем на 30% больше меда, чем с пасек, расположенных в глубине лесных массивов (Wellenstein, 1960).
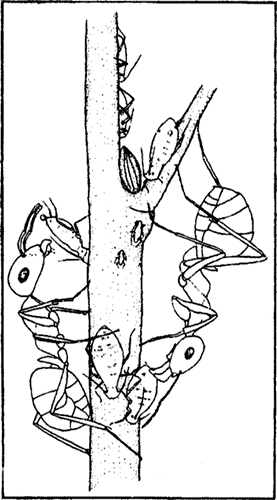
взаимовыгодные отношения между муравьями и выделяющими падь насекомыми: слизывая падь, муравьи охраняют этих насекомых.
(от греч. symbíosis — сожительство), в узком смысле под симбиозом понимают такое сожительство особей двух видов, при котором оба партнёра вступают в непосредственное взаимодействие с внешней средой; регуляция отношений с последней осуществляется совместно усилиями, сочетанной деятельностью обоих организмов (ср. Паразитизм). В широком смысле симбиоз охватывает все формы тесного сожительства организмов разных видов, включая и паразитизм, который в этом случае называется антагонистическим симбиозом
подотряд насекомых отряда равнокрылых. Длина 0,5-6 мм. Св. 2,5 тыс. видов. Распространены главным образом в умеренном поясе Северного полушария. Живут колониями на растениях. Многие тлевые опасные вредители, некоторые переносчики ряда вирусных заболеваний растений. Сахаристые испражнения тлевых (падь) привлекают муравьев, которые охраняют тлевых.
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
 Formica rufa Linnaeus, 1761 — рыжий лесной муравей
Formica rufa Linnaeus, 1761 — рыжий лесной муравейсладкая густая жидкость, выделяемая тлями, червецами и другими насекомыми, питающимися соком растений. Медоносная пчела собирает падь с листьев и перерабатывает в падевый мед.
общее название нескольких семейств насекомых подотряда кокцидовых. Св. 1600 видов, преимущественно в тропиках и субтропиках. У тропических лаковых червецов покровы образованы застывающими на воздухе лаковыми выделениями. Многие (напр., мучнистые червецы) вредители растений, особенно субтропических (напр., цитрусовых).
Устарел, исключен.
 Formica sanguinea Latreille 1798 — кровавый муравей-рабовладелец
Formica sanguinea Latreille 1798 — кровавый муравей-рабовладелецУстарел, исключен.
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
функциональное образование, состоящее из материнской и дочерних семей, поддерживающих регулярные родственные отношения
 Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky ant
Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky antнеобратимое историческое развитие живой природы. Определяется изменчивостью, наследственностью и естественным отбором организмов. Сопровождается приспособлением их к условиям существования, образованием и вымиранием видов, преобразованием биогеоценозов и биосферы в целом.
(от греч. mýrmēx - муравей и philía - любовь, склонность), использование муравьями особенностей строения или выделений некоторых растений. Муравьи селятся в полых стволах растений, колючках, междоузлиях или в сообщающихся ходах клубнеобразных стеблей. В других случаях муравьи питаются железистыми выделениями растений, содержащими белки, сахара и жиры.
Другой формой Мирмекофилия является приспособленность ряда животных к обитанию в гнёздах муравьев. Отношения муравьев к мирмекофилам могут быть нейтральными; нередки случаи симбиоза и комменсализма.
(от греч. oikos дом, жилище, местопребывание и ...логия), наука об отношениях живых организмов и образуемых ими сообществ между собой и с окружающей средой. Термин «экология» предложен в 1866 Э. Геккелем. Объектами экологии могут быть популяции организмов, виды, сообщества, экосистемы и биосфера в целом.
взрослая особь
сборное название насекомых подотряда жалящих перепончатокрылых (кроме пчел и муравьев). Длина до 40 мм (шершни). Ок. 23 тыс. видов. Распространены широко. Опылители растений. Личинки некоторых ос паразиты других насекомых.
надсемейство летающих насекомых отряда перепончатокрылых, родственное осам и муравьям. Существует около 20 тысяч видов пчёл. Распространены на всех континентах, кроме Антарктиды. Пчёлы питаются нектаром и пыльцой, используя нектар главным образом в качестве источника энергии, а пыльцу для получения белков и других питательных веществ.






