






Хотя строение гнезд даже в пределах одного вида муравьев сильно изменяется, все же возможно свести все их разнообразие к определенной системе типов. Попытки классификации гнезд этих насекомых предпринимались неоднократно. Наиболее известна система, предложенная А. Форелем (Forel, 1874, 1928) и сохранившаяся практически без изменений до настоящего времени (Длусский, 1967а, 19726; Sudd. 1967). Однако в этой классификации все гнезда, сделанные в земле и не имеющие наружных построек, относятся к одному недифференцированному классу. Некоторые авторы (Francoeur, Maldague, 1966; Delye, 1968) предлагали разделять гнезда, относящиеся к этому классу, по местообитаниям (гнезда на песчаной почве, на глинистой почве и т.д.), но к собственно классификации гнезд такое деление имеет весьма слабое отношение. Такой подход становится понятным, если учесть что классификация разрабатывалась в основном применительно к видам, обитающим в лесах умеренной зоны и влажных тропиков. В этих условиях многие виды строят гнезда с различными наружными постройками, гнезда в разлагающейся древесине, на деревьях и т.д., а устройство подземных гнезд без наружных построек не отличается ни сложностью, ни разнообразием. В то же время в пустынях гнезда муравьев, несмотря на все их многообразие, формально относятся к одному типу, поскольку лишь незначительное число видов обитает в древесине (Crematogaster, Tetraponera, немногие Camponotus) или возводит земляные холмики (некоторые муравьи-жнецы Сахары и Северной Америки).
Отсутствие попыток классификации подземных гнезд муравьев было связано и с недостатком материала. Правда, в аридных и семиаридных районах США было раскопано и описано довольно много гнезд Pogonomyrmex (Wray, 1938; Lavigne, 1969; Whitford et al., 1976) и отдельные гнезда Myrmecocystus (McCook, 1881; Creighton, Crandall, 1954) и Veromessor (Wheeller, Rissing, 1975a), а в семиаридных районах Австралии — Myrmecia (Gray, 1971b) и Iridomyrmex (Ettershank, 1968) и в Сахаре — Messor (Tohme, 1972) и нескольких других родов (Delye, 1968). Однако авторы приведенных работ (кроме Дели) имели дело лишь с одним-двумя видами. П. И. Мариковский (1957, 1962, 1979) приводит данные о строении гнезд у ряда видов муравьев пустынь Казахстана. Попытка провести сравнительный анализ гнездостроения пустынных муравьев была сделана Ж. Дели (Delye, 1968), который описывает гнезда 14 видов Сахары. Но этот автор не делал раскопок глубже 1 м, а гнезда пустынных муравьев обычно уходят на большую глубину и у многих видов достигают грунтовой воды. Первой попыткой дать классификацию типов гнезд у муравьев, обитающих в пустынях, была наша работа (Длусский, 1974а), основанная на данных полной раскопки гнезд 14 видов и частичной (до 1—1,5 м) раскопки 13 видов муравьев. После выхода в свет этой статьи был собран дополнительный материал, и в настоящее время мы располагаем данными полной раскопки 46 гнезд 21 вида и частичных раскопок большого числа гнезд 31 вида муравьев, сделанных в Кызылкумах, Каракумах и Каршинской «степи».
Прежде чем перейти к описанию типов гнезд, необходимо остановиться на терминологии. Элементарными структурными единицами любого подземного гнезда муравьев являются ходы (галереи) и камеры. Ходы, как правило, имеют в сечении форму круга. Диаметр их варьирует у разных видов от 1 до 10 мм. Иногда муравьи утрамбовывают стенки ходов и камер или пропитывают их выделениями желез. Камеры у большинства видов располагаются горизонтально и в плане обычно имеют форму вытянутого овала или круга. Пол камер, как правило, плоский и выглаженный, потолок либо параллелен полу, либо сводчатый; часто поверхность его имеет гранулированную структуру. У мелких муравьев часто камеры имеют неправильную форму. В этом случае личинки не лежат на полу камеры, как обычно, а подвешиваются к ее потолку и стенам. Помимо обычных камер, в которых находятся муравьи и расплод, у некоторых видов имеются камеры для хранения запасов (КЗ) /Приведенные в скобках сокращения использованы для обозначения структурных элементов гнезд на рисунках./ и камеры, заполненные отбросами (КО). Часто их строение не отличается от строения обычных камер. У Messor и Leptothorax semenovi камеры с запасами иногда имеют сферическую или близкую к ней форму. Камеры с отбросами особенно характерны для Messor и Cataglyphis. У первых они заполнены мякиной, а у вторых — хитиновыми остатками съеденных насекомых; в них обычно встречается много неспециализированных мирмекофилов. И в том и в другом случае обычно это большие плоские камеры, часто в плане неправильной формы, располагающиеся под самой поверхностью почвы. У американских пустынных муравьев, разводящих грибы (Attini), имеются специальные камеры для грибных садов (Wheeler, 1907b, 1910а, 1911). Иногда в гнездах муравьев выделяют «царскую» камеру, где находится самка. Обычно это самая глубокая и крупная камера. Однако у муравьев самки подвижны (за исключением редких у пустынных муравьев случаев физогастрии) и могут свободно передвигаться по гнезду. Весьма возможно, что их нахождение в самой глубокой камере объясняется тем, что они переходят в нее во время раскопки гнезда.
В гнездах пустынных муравьев можно выделить и более крупные структурные элементы, отличающиеся большим или меньшим постоянством (рис:52).
Вход (ВХ) в гнездо часто представляет собой просто отверстие. Иногда вокруг него заметны выбросы почвы. В тех случаях, когда муравьи, обладающие псаммофором, не трамбуют вынесенных комков, эти выбросы представляют собой шарики почвы (например, у Camponotus или Cataglyphis foreli). У некоторых видов вход окружен валиком почвы, который иногда (например, у Messor variabilis) бывает очень высоким и напоминает кратер. Многие пустынные виды на то время, когда рабочие не активны на поверхности, закрывают вход частичками почвы, мелкими камешками или сухими растительными остатками.
Поверхностная горизонтальная система ходов и камер (ПГС) представляет собой сеть камер в поверхностном слое почвы (0—40 см). Эти камеры часто соединены между собой ходами без видимого порядка, но иногда различается несколько неявственных ярусов. В камерах ПГС весной находится основная масса расплода.
Поверхностная горизонтальная галерея (ПГГ) у некоторых видов заменяет ПГС. В простейшем случае это горизонтальный ход, но иногда она расширена и скорее напоминает очень длинную камеру. В ряде случаев наблюдается ветвление этой галереи. Кроме того, при помощи поверхностных галерей соединяются различные отделы сложных гнезд муравьев.
Ствол (С) — вертикальный или наклонный ход, начинающийся от входа, поверхностной галереи или одной из камер ПГС. В него открываются все или почти все глубинные камеры гнезда. Ствол либо заканчивается слепо, либо в конце его имеется камера, иногда довольно крупная («царская» камера).
Глубинная горизонтальная система ходов и камер (ГГС) иногда строится в глубоком влажном горизонте. В ней могут размещаться личинки во время сухого периода. Чаще всего ГГС представляет собой горизонтальную галерею с немногими камерами.
Станции (СТ) — это небольшие дополнительные гнезда, состоящие чаще всего из одного ствола с несколькими камерами. Они соединяются с главным гнездом поверхностными или глубинными горизонтальными галереями или связаны с ним дорогами. В станциях обычно находятся только рабочие особи, но иногда имеется и некоторое количество личинок. Tapinoma и Crematogaster строят станции у корней растений и разводят в них корневых тлей, червецов или цикадок.
Из всех муравьев аридных и семиаридных областей самые примитивные гнезда были обнаружены нами у Alloformica aberrans (рис:53). Они устроены даже проще, чем гнезда австалийских Myrmecia, считающихся самыми примитивными из ныне живущих муравьев. Гнездо A. aberrans — это толстый, сравнительно короткий (30—50 см) вертикальный ствол, начинающийся прямо от входа или (на крутых склонах) соединенный с ним короткой горизонтальной галереей. Настоящих камер у этого вида нет, но иногда ствол расширяется или образует широкие слепые отростки, напоминающие камеры. В конце ствол расширяется, образуя некоторое подобие выводковой камеры некоторых одиночных ос. Это расширение бывает плотно забито муравьями и расплодом. Гнезда австралийской пустынной Myrmecia dispar (Gray, 1971), как и гнезда A. aberrans, имеют толстый и сравнительно неглубокий (15—77 см) вертикальный ствол, соединяющийся непосредственно со входом. Однако у этого муравья имеются камеры, которые, правда, больше напоминают (за исключением главной камеры) короткие горизонтальные ходы. Заканчивается ствол главной камерой, в которой находится большая часть расплода. Число камер редко превышает 10 и лишь у наиболее многочисленной семьи (368 особей, т.е. несколько больше, чем у A. aberrans) достигало 15. Гнезда типа Alloformica и Myrmecia, во многом сходные с гнездами одиночных ос, можно отнести к сфекоидному типу гнезд муравьев.
Среди собственно пустынных муравьев Палеарктики (A. aberrans в настоящих пустынях не встречается) самым примитивным является гнездо, состоящее из вертикального ствола, с которым соединяются настоящие камеры, и не имеющее развитой ПГС. Иногда ствол не соединяется непосредственно со входом, а открывается в поверхностную галерею. В некоторых случаях эта галерея может ветвиться, иногда расширяется и имеет вид длинной плоской камеры (C. foreli), иногда с ней соединяется несколько камер. Подобные гнезда характерны для обитающих в Сахаре Cataglyphis emmae и C. albicans (Delye, 1968) и для C. emeryi (рис:54), C. pallida (рис:52), C. foreli (рис:55), C. cinnamomea (рис:56), Proformica epinotalis (рис:57) и Leptothorax semenovi (рис:58) Средней Азии. Численность населения в семьях этих видов редко достигает 1000 особей. Аналогичные гнезда встречаются также у молодых семей ![]() Cataglyphis aenescens (рис:59), C. setipes, Camponotus, Messor, т.е. видов, для которых характерны гнезда более сложного строения.
Cataglyphis aenescens (рис:59), C. setipes, Camponotus, Messor, т.е. видов, для которых характерны гнезда более сложного строения.
Наиболее распространенным типом гнезд у пустынных муравьев являются гнезда с развитой ПГС и одним или несколькими стволами. Такие гнезда встречаются у Cataglyphis bombycina Сахары (Delye, 1968), C. setipes (рис:60, рис:61) и ![]() C. aenescens (рис:59) пустынь Средней Азии, семьи которых насчитывают несколько тысяч особей. Они характерны для большинства видов Messor Сахары (Delye, 1968, 1971; Tohme, 1972), Южной Европы (Eidmann, 1926) и Средней Азии (Мариковский, 1957, 1962; Сапарлиев. 1972; Длусский, 1974) (рис:62), Myrmecocystus (МсСооk, 1881), Pogonomyrmex (Lavigne, 1969) и Veromessor (Wheeler, Rissing, 1975) аридных областей Северной Америки, Acantholepis semenovi, Camponotus turcestanus, C. fedtschenkoi пустынь Средней Азии и многих видов гумидной зоны (Длусский, 1967а). Такой же тип строения имеют гнезда молодых или слабых семей Camponotus xerxes, Tetramorium, Plagiolepis и др.
C. aenescens (рис:59) пустынь Средней Азии, семьи которых насчитывают несколько тысяч особей. Они характерны для большинства видов Messor Сахары (Delye, 1968, 1971; Tohme, 1972), Южной Европы (Eidmann, 1926) и Средней Азии (Мариковский, 1957, 1962; Сапарлиев. 1972; Длусский, 1974) (рис:62), Myrmecocystus (МсСооk, 1881), Pogonomyrmex (Lavigne, 1969) и Veromessor (Wheeler, Rissing, 1975) аридных областей Северной Америки, Acantholepis semenovi, Camponotus turcestanus, C. fedtschenkoi пустынь Средней Азии и многих видов гумидной зоны (Длусский, 1967а). Такой же тип строения имеют гнезда молодых или слабых семей Camponotus xerxes, Tetramorium, Plagiolepis и др.
Гнезда молодых семей всегда имеют один ствол. По достижении определенного порогового размера семья строит новый ствол. Этот порог будет весьма различным для разных видов: у Pogonomyrmex occidentalis он составляет около 700 особей (Lavigne, 1969), у Camponotus xerxes — 600—1000 особей, у Cataglyphis setipes — около 2,5 тыс. особей (в пересчете на осеннюю численность) (Длусский, 1974а). Максимальное число стволов также специфично для вида. Так, у ![]() C. aenescens в Средней Азии даже у самых сильных семей мы редко находили гнезда с двумя стволами, хотя для степной зоны описаны гнезда с пятью стволами (Резникова и др., 1977). У C. setipes число стволов может достигать трех, у Pogonomyrmex occidentalis (Lavigne, 1969), Messor aralocaspius и M. denticulatus — пяти, у M. intermedius — более десятка. Иногда со стволами соединяется глубинная горизонтальная система, расположенная в горизонте, увлажняемом капиллярной водой. Все стволы таких гнезд имеют в принципе одинаковое строение и несут одни и те же функции, поскольку мы всегда находим при раскопках в камерах каждого ствола и расплод, и (у муравьев-жнецов) запасы семян. У Cataglyphis, как правило, имеется всего один вход в гнездо, независимо от числа стволов. У Camponotus число входов приблизительно соответствует числу стволов. У Messor количество входов в гнездо не связано с числом стволов; входы в гнездо у этих муравьев не фиксированы и в зависимости от потребностей семьи могут открываться из разных поверхностных камер.
C. aenescens в Средней Азии даже у самых сильных семей мы редко находили гнезда с двумя стволами, хотя для степной зоны описаны гнезда с пятью стволами (Резникова и др., 1977). У C. setipes число стволов может достигать трех, у Pogonomyrmex occidentalis (Lavigne, 1969), Messor aralocaspius и M. denticulatus — пяти, у M. intermedius — более десятка. Иногда со стволами соединяется глубинная горизонтальная система, расположенная в горизонте, увлажняемом капиллярной водой. Все стволы таких гнезд имеют в принципе одинаковое строение и несут одни и те же функции, поскольку мы всегда находим при раскопках в камерах каждого ствола и расплод, и (у муравьев-жнецов) запасы семян. У Cataglyphis, как правило, имеется всего один вход в гнездо, независимо от числа стволов. У Camponotus число входов приблизительно соответствует числу стволов. У Messor количество входов в гнездо не связано с числом стволов; входы в гнездо у этих муравьев не фиксированы и в зависимости от потребностей семьи могут открываться из разных поверхностных камер.
Наиболее сложные односекционные гнезда обнаружены у Messor variabilis (Длусский, 1974). Стволы этих гнезд имеют разное строение и несут различные функции. В гнезде M. variabilis №65-145 (рис:63) в главном стволе и в камерах ПГС находились только небольшое количество рабочих и (в глубине ствола) запасы семян. Непосредственно под кратером кольцом располагались большие камеры, забитые мякиной. Весной в ПГС главного ствола находятся крылатые половые особи, готовые к вылету. От главного ствола на глубине 75 и 150 см отходили две горизонтальные галереи длиной 80 и 90 см с немногими камерами. В камерах нижней галереи находились основные запасы семян. Эти галереи соединяли главный ствол с двумя дополнительными (А и В), которые подходили к самой поверхности почвы, но не имели самостоятельных входов. Камеры одного из стволов были пустыми, а в камерах другого находился расплод (табл.22). Судя по тому, что при частичных (до 1—1,5 м) раскопках гнезд M. excursionis и M. lamellicornis мы ни разу не обнаружили в ПГС и главном стволе ни расплода, ни сколько-нибудь значительных запасов, можно ожидать, что подобное строение гнезд окажется и у этих видов.
Переход к такому специализированному строению гнезда наблюдается у ![]() Monomorium gracillimum. Так, в гнезде №71-22 (рис:64) от одного из двух главных стволов отходила горизонтальная галерея, которая заканчивалась дополнительным стволом с несколькими камерами, не имеющими самостоятельного входа. Запасы семян располагались в камерах ПГС, непосредственно соединяющихся с главным входом, а самка — в одной из камер дополнительного ствола. Однако расплод был обнаружен в камерах главных и дополнительного стволов и в глубинной горизонтальной системе. Молодые семьи этого вида имеют простые гнезда с ПГС и одним стволом.
Monomorium gracillimum. Так, в гнезде №71-22 (рис:64) от одного из двух главных стволов отходила горизонтальная галерея, которая заканчивалась дополнительным стволом с несколькими камерами, не имеющими самостоятельного входа. Запасы семян располагались в камерах ПГС, непосредственно соединяющихся с главным входом, а самка — в одной из камер дополнительного ствола. Однако расплод был обнаружен в камерах главных и дополнительного стволов и в глубинной горизонтальной системе. Молодые семьи этого вида имеют простые гнезда с ПГС и одним стволом.
На базе ПГС у некоторых видов пустынных муравьев формируются специальные структурные образования, среди которых следует отметить сушильные галереи Cataglyphis foreli и систему тоннелей, встречающуюся у многих видов. При раскопке гнезд C. foreli мы обнаружили, что ПГС их гнезд образуется уже упоминавшейся выше широкой и плоской поверхностной галереей. Такие слепо оканчивающиеся галереи имеют длину 1—2 м. Так, у гнезда №65—146 (рис:55) эта галерея имела длину 180 см, начиналась на глубине 10 см и заканчивалась на глубине 4 см, т.е. под самой поверхностью почвы. При высоте около 2 см галерея местами имела ширину 15 см. В конце галереи находились трупы термитов, среди которых были как свежие, так и совершенно высохшие. Интересно, что при раскопке этого и других гнезд C. foreli в камерах с личинками мы находили только высушенный корм (чаще всего термитов). У видов рода Monomorium от ПГС гнезда отходят поверхностные тоннели до 2 м длиной. В конце каждого такого тоннеля имеется вход (рис:64). Под входом имеется либо одна-две камеры, либо просто тоннель перед входом несколько расширяется. Иногда тоннели соединяют гнездо с подземными камерами, в которых находятся корневые тли, червецы или цикадки. Система тоннелей характерна не только для пустынных видов и описана у многих муравьев умеренной зоны (Brian, Downing, 1958; Захаров, 1972).
При увеличении размеров семьи может наступить момент, когда гнездо, даже имеющее несколько стволов, не может вместить все население. Одним из путей ликвидации этой диспропорции является образование колоний (Захаров, 1972). Однако образование колоний возможно лишь при полигинии. У моногинных и некоторых олигогинных видов распространен другой путь — образование многосекционных гнезд (Длусский, 1974а). На некотором расстоянии от старого гнезда строится новое гнездо, соединенное с первым, затем второе, третье и т.д. Эти гнезда, сохраняющие постоянную связь между собой через подземные галереи, мы назвали секциями. Исследования А. А. Захарова (1976) подтвердили наше предположение, что население одной секции представляет собой колонну (подробнее см. ниже). В простейшем случае многосекционное гнездо — это цепочка секций, соединенных подземными галереями. Именно такой тип гнезда часто наблюдается у Tapinoma karavaievi, Tetramorium schneideri (рис:65), Camponotus turcestanicus, Pheidole arenarum и Plagiolepis. В случае олигогинии, как, например, у P. pallescens (рис:66), самки более или менее равномерно распределяются между секциями. Линейное расположение секций в гнезде не является обязательным. У T. schneideri, например, мы находили гнезда с кольцевым и сетчатым расположением секций. Многосекционные гнезда характерны и для некоторых видов, обитающих в гумидных областях, в частности Formica picea и F. cinerea.
В случае моногинии самка располагается в одной из секций многосекционного гнезда, что является предпосылкой для возникновения функциональной дифференциации секций. Такая функциональная дифференциация встречается у Camponotus xerxes. Крупные семьи этого вида обитают в гнездах, насчитывающих до десятка секций и более 20 входов. Такое гнездо может занимать площадь более 15 м². Одно такое гнездо (№17-11) было раскопано в Акыбае (рис:67). Гнездо состояло из семи секций и имело девять входов. Две секции (КЛ и М) не были вскрыты, а в секции ЗИ были подсчитаны только рабочие особи (448 экз.). Данные о численности населения и его составе в остальных четырех секциях приведены в табл.23. Как видно из блок-схемы, каждая секция имела один — три ствола, открывавшихся в общую ПГС глубиной до 40 см. ПГС разных секций соединялись между собой галереями, проходившими на глубине 5—30 см. Число стволов в секциях соответствовало числу входов, однако прямо связать входы со стволами можно не всегда (например, входы А, Б и В стволы la, 1b и 1с). Кроме того, имелась одна секция (6) с единственным стволом, не имеющая самостоятельного входа. В ней находилась основная масса расплода и единственная самка с функционирующими яичниками. Две другие самки имели яичники, не отличавшиеся от таковых у крылатых самок, и размещались в вертикальном стволе 1с и в ПГС, связанной со входом Ж.
На основании данных 1965—1971 гг. мы описывали гнездо Monomorium barbatulum как односекционное с функциональной дифференциацией стволов и системой станций (Длусский, 1974а, 1975а). Однако раскопки 1974 г. заставили пересмотреть эту точку зрения. Гнезда этого крошечного (около 3 мм) муравья — самые трудные для раскапывания гнезда муравьев, с которыми нам приходилось сталкиваться. Небольшие камеры рассеяны на большом пространстве и соединяются длинными ходами диаметром менее 1 мм, часто проходящими в сухом песке. Неосторожное движение приводит к тому, что песок оплывает и засыпает ход, после чего раскопки можно прекращать. Наиболее полно вскрытое в 1971 г. гнездо (№71—28, рис:68/2/) имело три ствола, соединявшиеся горизонтальными галереями, проходившими под самой поверхностью почвы на глубине 1—2 см. Два ствола находились рядом и имели общую ПГС с одним входом. В одном из стволов находилась самка, 260 личинок и 27 куколок. В этом стволе и в ПГС была сосредоточена основная масса рабочих (344 экз.). Во втором стволе было всего 20 рабочих, расплода не было, а в камерах было обнаружено значительное количество семян. От этого ствола отходила на глубине 117 см горизонтальная галерея длиной 217 см с четырьмя камерами, которая заканчивалась слепо. В ней находилось 10 рабочих и не было ни расплода, ни запасов семян. В метре от входа в ПГС этой секции находилась еще одна секция из одного ствола, имеющая самостоятельный вход, которую мы приняли за станцию. В ней находилось 55 рабочих, 24 личинки и 9,6 мг семян. На глубине 50 см ствол либо кончался, либо был потерян. С главной секцией этот ствол соединялся тонкой галереей, идущей под самой поверхностью почвы. В том же году вскрывали еще несколько гнезд, но удалось раскопать не больше одного ствола и ни самок, ни большого количества расплода в этих гнездах не было обнаружено. Когда ствол удавалось проследить до глубины около 1 м, всегда обнаруживалась горизонтальная глубинная галерея. В одном случае удалось проследить ее на расстоянии 195 см, но затем ход был потерян.
В 1974 г. в окрестностях Карши нам удалось проследить горизонтальную глубинную галерею M. barbatulum на расстоянии 285 см. В обоих направлениях галерея продолжалась дальше, но ход был потерян. Было обнаружено, что с галереей соединялись две секции, каждая из которых состояла из ПГС и одного ствола (рис:68/1/). Входы в эти секции находились на расстоянии 125 см друг от друга. Полностью вскрыть большое гнездо M. barbatulum нам ни разу не удалось, но на основании приведенных данных, а также данных картирования входов на площадках (рис:4) можно составить более или менее полную картину. Очевидно, у этого муравья имеются многосекционные гнезда, занимающие площадь несколько квадратных метров (иногда более 10 м²). Большинство секций содержит только рабочих, некоторое количество расплода и запасы семян, а самка и основная масса расплода находятся лишь в одной секции. Отдельные секции соединяются между собой длинными галереями, проходящими либо под самой поверхностью почвы (в песчаной пустыне, где обитают M. barbatulum, они должны летом разрушаться), либо на глубине от 50 (гнездо №74-19) до 135 см (гнездо №71-23).
На основании всех приведенных данных можно предположить следующую классификацию подземных гнезд муравьев, обитающих в аридных областях:
I. Сфекоидный тип (гнезда, не имеющие настоящих камер).
II. Формикоидный тип (гнезда с настоящими камерами, соединенными ходами).
А. Односекционные гнезда:
1) без развитой ПГС, с одним стволом;
2) с развитой ПГС и одним или несколькими стволами:
а) без функциональной дифференциации стволов;
б) с функциональной дифференциацией стволов.
1) без функциональной дифференциации секций;
2) с функциональной дифференциацией секций.
Связь между этими типами и адаптивное значение гнезда в целом и его отдельных структур будут рассмотрены в следующем разделе.
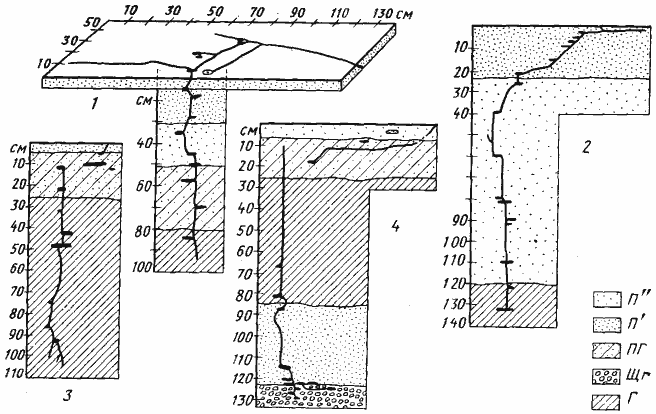
Рис. 52. Гнезда Cataglyphis pallida
1 — Акыбай, №71-10, 2 апреля 1971 г.; 2 — там же, №71-08, 2 апреля 1971 г.; 3 — Дарья-сай, Юго-Западные Кызылкумы, № 65-140; 4 — там же, №65-150, 4 июня 1965 г. Обозначения на рисунках 52 —68: В — горизонт почвы с грунтовой водой; Вх — вход в гнездо; Г — глина; Гг — глина с большим содержанием гипса; Гп — кристаллы гипса; ГГС — глубинная система горизонтальных галерей и камер; КО — камеры с остатками пищи (хитин насекомых, мякина); КТ — камеры с колониями тлей на корнях; Л — лёсс; Л'' — сухой лёсс; Лг — лёсс с большим содержанием гипса; ОС — остатки хитина и старые коконы, захороненные в почве; П' — влажный песок; П'' — сухой песок; ПГ — тяжелая супесь или легкий суглинок; ПГС — поверхностная система горизонтальных галерей и камер; РО — почва с большим количеством захороненных растительных остатков; С — ствол гнезда; Щ — щебень или галька; Щг — щебень или галька, сцементированные гипсом; O — яйца; L — личинки; P — куколки; r — плерэргаты; S — запасы семян; ♂ — рабочие (обычно не указываются); ♀ —бескрылые самки; ᴓ — диаметр хода или галереи в мм
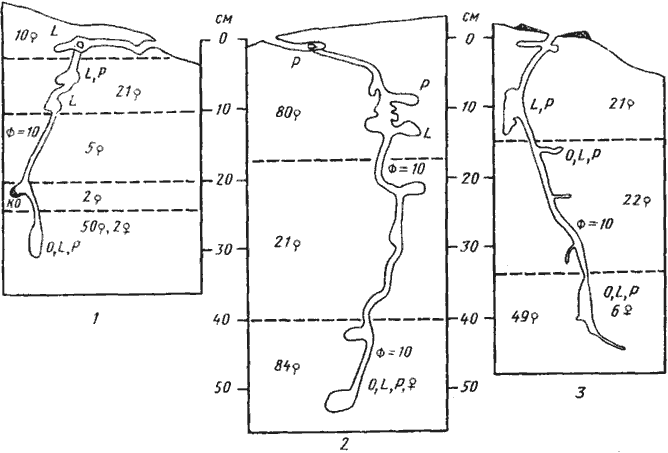
Рис. 53. Гнезда Alloformica aberrans
1 — Варганза, № 78-170, 28 мая 1978 г.; 2 — там же, № 78-169, 29 мая 1978 г.; 3 — там же, № 78-169а (только что выстроенное гнездо), 29 мая 1978 г.
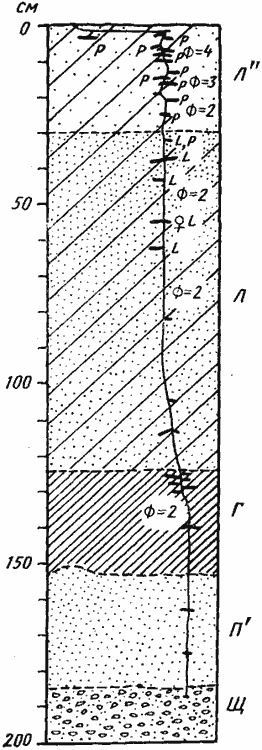
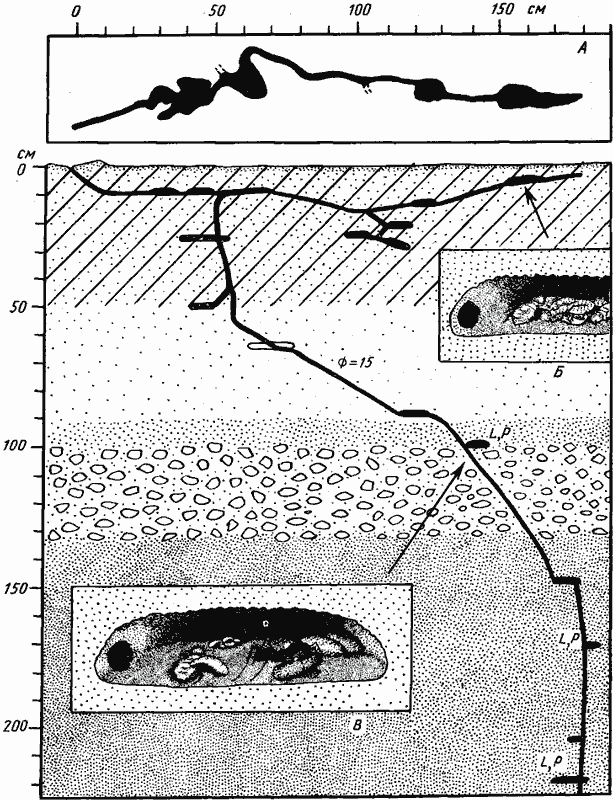
Рис. 55. Гнездо Cataglyphis foreli
(Дарья-сай, Юго-западные Кызылкумы, № 65—146, 29 мая 1965 г.)
А — поверхностная горизонтальная галерея, вид сверху; Б — камера с запасом термитов; В — камера с расплодом; на переднем плане личинка, поедающая высушенного термита. Раскопка была сделана только до глубины 220 см, но ствол уходил глубже
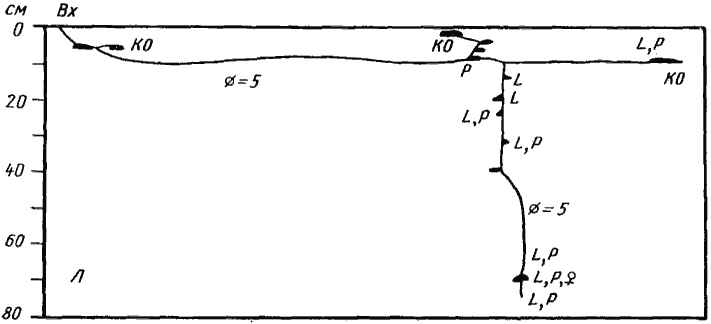
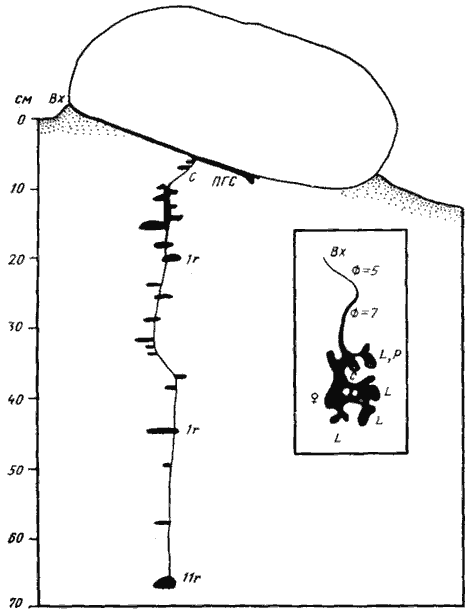
Рис. 57. Гнездо Proformica epinotalis
(Варганза, № 78-200, 1 июня 1978 г., лёсс со щебнем)
Гнездо под камнем; на врезке — поверхностная горизонтальная система сверху
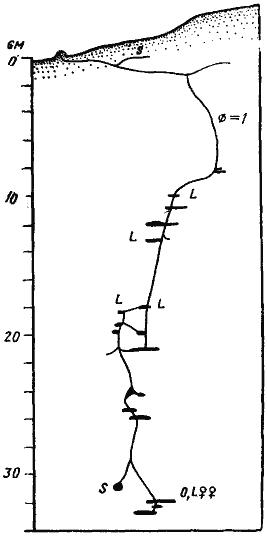
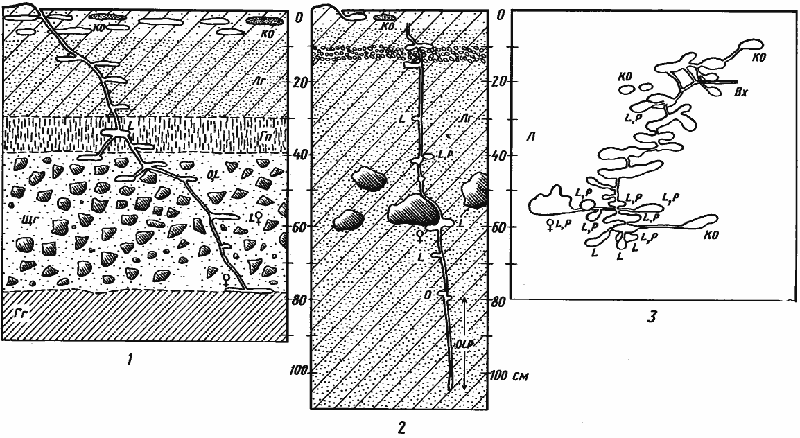
Рис. 59. Гнезда
1 — Аяк-Гужумды, № 65-116, 7 мая 1965 г., солончак; 2 — там же, № 65-118, 12 мая 1965 г.; вода на глубине 130 см; горизонтальная поверхностная система на рисунке, не изображена, так как уходит перпендикулярно плоскости рисунка; ее самая удаленная камера в 70 см от входа; 3 — Варганза, № 78-201, 30 мая 1978 г.; гнездо располагалось на крутом лёссовом обрыве, так что вход находился ниже ПГС
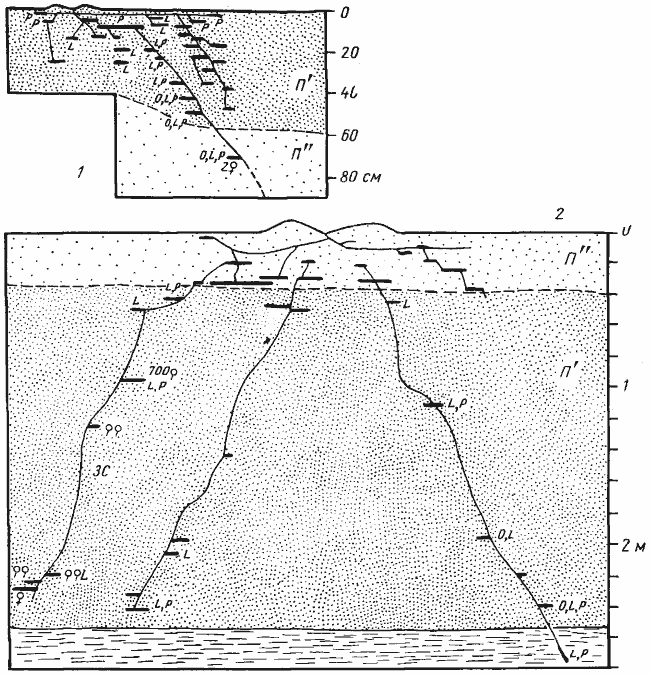
Рис. 60. Односекционные гнезда Cataglyphis setipes
(Акыбай)
1 — 15 мая 1971 г.; ствол засыпан и вся жизнь семьи была сосредоточена в верхнем влажном горизонте почвы; 2 — 1 сентября 1971 г.; ЗС — ствол, камеры которого были забиты рабочими, очевидно уже подготовленными к зимовке
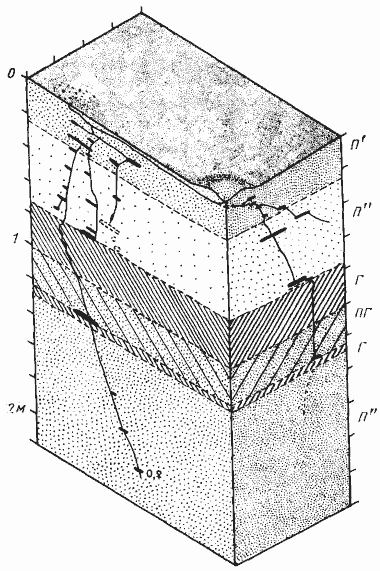
Рис. 61. Необычное двухсекционное гнездо Cataglyphis setipes
(Акыбай, 25—29 марта (971 г.)
Муравьи только что вышли из зимовки и в гнезде было только небольшое количество яиц, а личинки и куколки отсутствовали
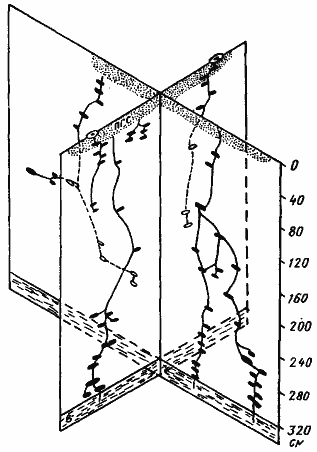
Рис. 62. Гнездо Messor aralocaspius с четырьмя стволами
(Акыбай, № 71-39, 28 апреля — 1 мая 1971 г.) [Длусский, 1974а]
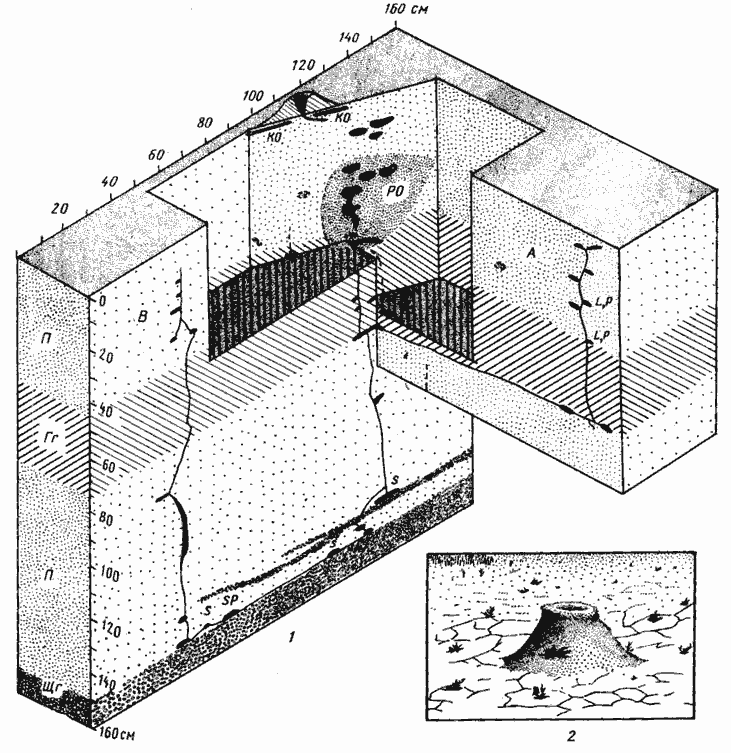
Рис. 63. Гнезда Messor variabilis
1 — Дарья-сай, № 65-145, 27-29 мая 1965 г.; пунктиром обозначены заброшенные камеры и ходы забитые разложившимися органическими остатками; 2 — вход в гнездо Messor variabilis (на такыре, Акыбай)
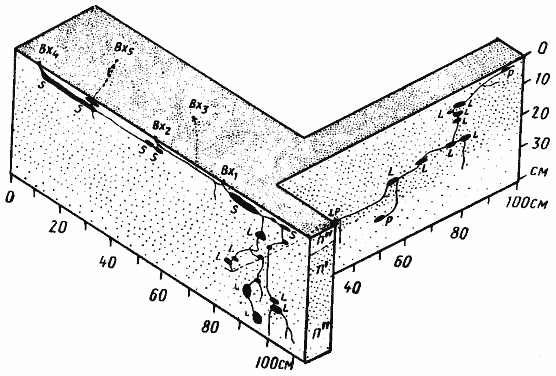
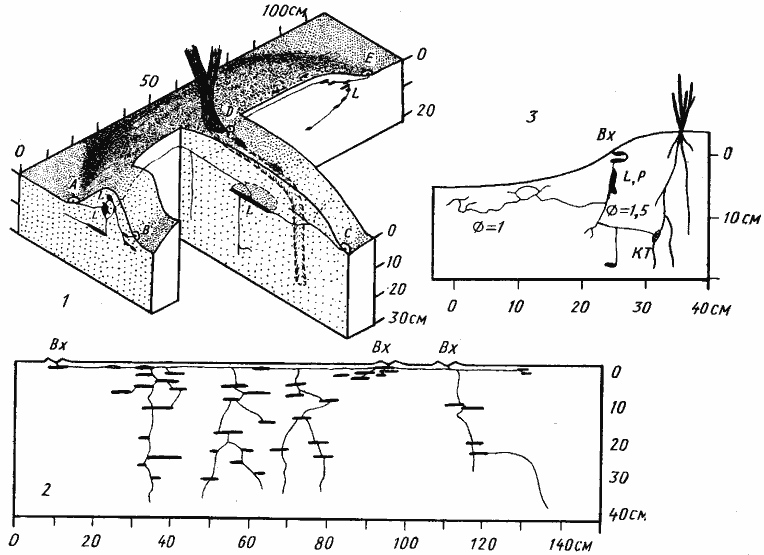
Рис. 65. Гнезда Tetramorium
1 — диффузное гнездо T. schneideri с тремя секциями (входы АВ, CD и Е), Акыбай, № 71-07, 1 апреля 1971 г., в семье около 500 рабочих; 2 — компактное гнездо того же вида с четырьмя линейно расположенными секциями и тремя входами, Акыбай, № 71-20, 24 апреля 1971 г., 2300 рабочих; 3 — небольшое гнездо T. inerme, Акыбай, № 71-09, 2 апреля 1971 г.
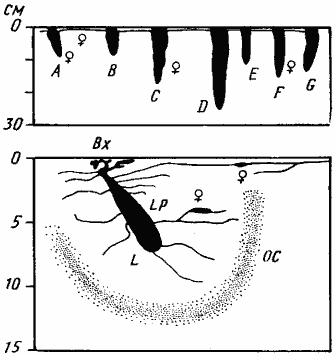
Рис. 66. Гнездо Plagiolepis pallescens
(Акыбай, № 71-05, 29 марта 1971 г.)
Вверху — схема расположения секций, внизу — секция А
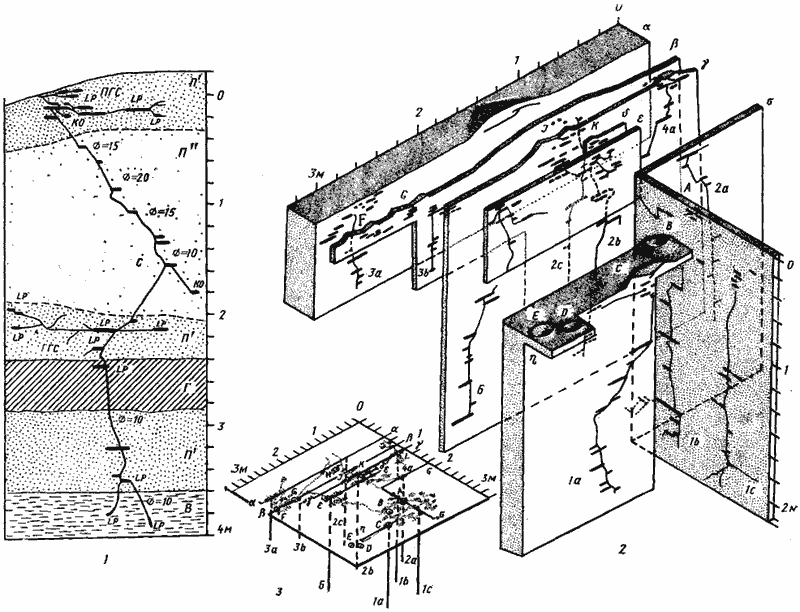
Рис. 67. Два гнезда Camponotus xerxes
1 — Акыбай, № 71-19, 21—22 апреля 1971 г., молодая семья с 859 рабочими; 2, 3 — Акыбай, № 71-11, 8—17 апреля 1971 г. Гнездо с населением около 6 тыс. рабочих, имевшее 6 секций, 11 входов и 10 стволов в пяти раскопанных секциях (секция 5 с входами D и Е не раскапывалась): 2 — изображены те срезы через гнездо, на которых обнажались стволы. Глубина стволов 1а и 1с была соответственно 220 и 240 см. Поскольку при раскопке этих стволов были вскрыты жилы грунтовой воды, вода в яме поднялась вначале до уровня 200 см, а затем — 180 см и поэтому остальные стволы вскрыть до конца не удалось; 3 — это же гнездо изображено схематически. На схеме показано взаимное расположение разрезов, поверхностная горизонтальная система (точками) и стволы гнезда (вертикальными линиями). Разрезы обозначены буквами греческого алфавита, входы в гнездо — заглавными латинскими буквами и стволы — буквенно-цифровым индексом, где цифра означает номер секции. Секция 6 не имела самостоятельного входа и в ней находились самка и основная масса расплода
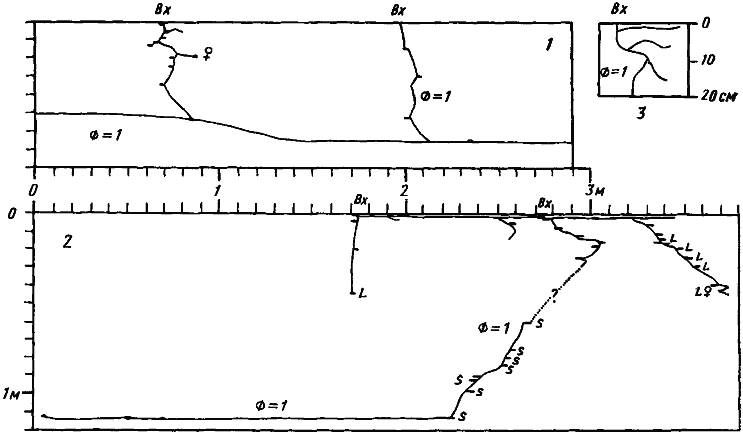
Рис. 68. Гнезда Monomorium barbatulum
1 — фрагмент гнезда № 74-19, Карши, 25 мая 1974 г., лессовидный суглинок, до 30 см сухой, глубже — влажный: 2 — гнездо № 71-28, Акыбай, 6 мая 1971 г., песок, влажный сверху и сухой на глубине более 50 см; 3 — система тонких поверхностных галерей в сухом песке гнезда № 71-06, Акыбай. 29 марта 1971
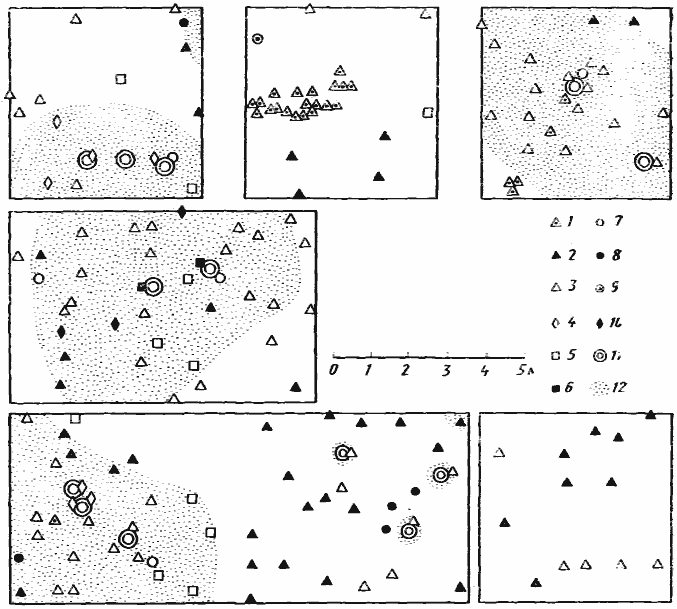
Рис. 4. Размещение входов в гнезда муравьев на шести пробных площадках в саксауловом лесу
Акыбай, 2 мая 1971 г.)
1 — Monomorium kusnezovi;
2 — M. barbatulum;
3 —
4 — Pheidole pallidula;
5 — Tetramorium inerme;
6 — Crematogaster subdentata;
7 — Plagiolepis pallescens;
8 — Acantholepis semenovi;
9 — Cataglyphis pallida;
10 — Cardiocondyla uljanini;
11 — саксаулы;
12 — подстилка под саксаулами
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
Всего 427 видов.
Всего насчитывает 85 видов.
Всего 965 видов.
Всего 60 видов.
Всего 30 видов.
Всего 90 видов.
Всего 58 видов.
Всего 106 видов.
Старое название: Leptothorax semenovi
Всего 65 видов.
полисахарид, образованный остатками аминосахара ацетилглюкозамина. Основной компонент наружного скелета (кутикулы) насекомых, ракообразных и других членистоногих. У грибов заменяет целлюлозу, с которой сходен по химическим и физическим свойствам и биологической роли.
(от греч. mýrmēx - муравей и philía - любовь, склонность), использование муравьями особенностей строения или выделений некоторых растений. Муравьи селятся в полых стволах растений, колючках, междоузлиях или в сообщающихся ходах клубнеобразных стеблей. В других случаях муравьи питаются железистыми выделениями растений, содержащими белки, сахара и жиры.
Другой формой Мирмекофилия является приспособленность ряда животных к обитанию в гнёздах муравьев. Отношения муравьев к мирмекофилам могут быть нейтральными; нередки случаи симбиоза и комменсализма.
сильное вздутие/увеличение брюшка. Физогастрическая самка имеет гипертрофированные яичники, благодаря чему отличается огромной продуктивностью. Встречается у моногинных видов муравьев.
специальное приспособление для переноски сухой почвы, распространенн у пустынных муравьев, образован длинными отстоящими волосками (аммохетами), часто имеющими форму латинской буквы J, расположен на нижней поверхности головы.
ксеробионт
земляной вал при выходе из жилого муравейника (секции), не имеющего наземного купола.
Всего 62 вида.
подотряд насекомых отряда равнокрылых. Длина 0,5-6 мм. Св. 2,5 тыс. видов. Распространены главным образом в умеренном поясе Северного полушария. Живут колониями на растениях. Многие тлевые опасные вредители, некоторые переносчики ряда вирусных заболеваний растений. Сахаристые испражнения тлевых (падь) привлекают муравьев, которые охраняют тлевых.
общее название нескольких семейств насекомых подотряда кокцидовых. Св. 1600 видов, преимущественно в тропиках и субтропиках. У тропических лаковых червецов покровы образованы застывающими на воздухе лаковыми выделениями. Многие (напр., мучнистые червецы) вредители растений, особенно субтропических (напр., цитрусовых).
сборное название насекомых подотряда жалящих перепончатокрылых (кроме пчел и муравьев). Длина до 40 мм (шершни). Ок. 23 тыс. видов. Распространены широко. Опылители растений. Личинки некоторых ос паразиты других насекомых.
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
Всего 4 вида.
ксеробионт
Старое название: Proformica deserta
Старое название: Cataglyphis cinnamomea
 Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунок
Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунокгигрофил, галобионт
Старое название: Cataglyphis flavigastra
ксеробионт
Старое название: Cataglyphis bombycina
Старые названия: Acantholepis semenovi, Acantholepis transcaspiensis
Всего 415 видов
Всего 60 видов.
ксеробионт
гигрофил
гнездо, состоящее из повторяющихся стереотипных блоков (секций), каждый из которых включает все видоспецифичные элементы (ствол, камеры и т. д.)
 Monomorium destructor (Jerdon, 1851) — изящный муравей, муравей-разрушитель
Monomorium destructor (Jerdon, 1851) — изящный муравей, муравей-разрушительСтарое название: Monomorium gracillimum
отряд общественных насекомых. Общины, разделенные на касты, состоят из крылатых и бескрылых особей. Строят подземные и наземные (до 15 м высотой) гнезда (термитники). Ок. 2600 видов, главным образом в тропиках; в России 2 вида: один в районе Сочи, второй, вероятно, завезенный из Японии или Китая, во Владивостоке. Многоядны. Разрушают древесину, портят бумагу, кожу, сельскохозяйственные продукты и др. Термиты, обитающие в почве, полезны как почвообразователи.
функциональное образование, состоящее из материнской и дочерних семей, поддерживающих регулярные родственные отношения
наличие в семье нескольких яйцекладущих самок. Смотри: моногиния, олигогиния.
наличие в семье одной яйцекладущей самки. Смотри: полигиния, олигогиния.
ситуация когда в гнезде живут несколько цариц, которые «не терпят» друг друга и занимают внутри муравейника отдельные территории. Смотри: моногиния, полигиния.
организационно оформленная мультифункциональная часть семьи (подсистема), имеющая постоянный состав и способная перейти к самостоятельному существованию.
гигрофил, галобионт
Старое название: Pheidole arenarum Ruzsky
олигогинны
рабочие с сильно раздутым брюшком; это каста, так называемых, "медовых бочек".
форма поликалии, при которой семья рассредоточена по готовым камерам и небольшим гнездам, не имеющим постоянного состава. Функциональной дифференциации таких гнезд и камер нет.
галобионт
Subspecies: Monomorium indicum kusnezowi Santschi, 1928






